Caracterização de cultivares portuguesas de Castanea
sativa Mill.: uma análise proteómica
Dissertação de Mestrado em
Genética Molecular Comparativa e Tecnológica
Michael Gaspar Freitas
Professor Doutor Gilberto Igrejas
Professor Doutor Hugo Santos
Presidente:___________________________________________________ 1º Vogal:____________________________________________________ 2º Vogal:____________________________________________________ Orientador:____________________________________________________ Classificação:_________________________________________________ Data: _________/_________/_________
Orientador
__________________________________________________ Professor Doutor Gilberto Paulo Peixoto Igrejas
Departamento de Genética e Biotecnologia Universidade de Trás-os-Montes e Alto Douro
Co-orientador
__________________________________________________ Professor Doutor Hugo Santos
Departamento de Química, Bioscope Group Universidade Nova de Lisboa
Ao terminar esta dissertação, desejo expressar o meu sincero reconhecimento pelo apoio e pela contribuição de diversas pessoas que tornaram possível a realização da mesma.
À Universidade de Trás-os-Montes e Alto Douro, na pessoa do magnífico Reitor Professor Doutor António Fontainhas Fernandes, que me acolheu para a realização desta dissertação, o meu sincero agradecimento pelos meios disponibilizados.
À direcção do mestrado, pela oportunidade de frequentar o mesmo e pela disponibilidade e prontidão que sempre demonstrou na resolução de qualquer dúvida ou problema que pudesse surgir.
Especialmente ao Professor Doutor Gilberto Igrejas, pela oportunidade de me ter orientado neste trabalho, por todo o interesse e empenho demonstrado no meu percurso académico, pela sua amizade e transmissão de inúmeros ensinamentos. Um muito Obrigado.
Ao Doutor Hugo Santos da Universidade Nova de Lisboa, por ter aceite ser meu co-orientador, por me ter recebido na Costa da Caparica e transmitido todos os ensinamentos sobre espectrometria de massa. Agradeço igualmente ao Professor Doutor José Luís Capelo Martinez, por me ter acolhido na Universidade Nova de Lisboa e no Grupo Bioscope, e por toda sua hospitalidade, tendo sido essencial para a realização deste trabalho.
Agradeço também ao Professor Doutor Jorge Ventura pelo fornecimento das amostras utilizadas neste trabalho.
A todos os meus colegas de laboratório, com um especial destaque para o Tiago e Catarina que partilharam muitos momentos deste trabalho, sendo fortes companheiros. Ao Eduardo e Susana pela paciência e disponibilidade de me instruírem em novas metodologias. E por fim ao Miguel, Luís, Alexandre e Cátia que foram verdadeiros amigos, sempre prontos a ajudar, ensinar, aconselhar e proporcionando também momentos de descontracção tão necessários quando o stress aumentava.
A todos os meus amigos de Lisboa, por me terem feito sentir em casa durante a minha estadia em Lisboa.
Agradeço aos meus pais pelo apoio que me deram ao longo de todo este processo, pela minha formação académica e como ser humano, pelo incentivo, conselhos e por toda a ajuda que
A todos os meus amigos, pela sua presença nesta fase da minha vida, que sempre me apoiaram. Finalmente a todos aqueles, que de uma forma directa ou indirecta me incentivaram e ajudaram na realização deste trabalho.
O castanheiro europeu (Castanea sativa Mill.) é uma espécie agro-florestal com elevada importância económica para as regiões Norte e Centro de Portugal. Durante muitos séculos, constituiu a base alimentar das populações rurais, devido à sua boa adaptação às condições edafoclimáticas, até ao aparecimento do milho e da batata.
As cultivares portuguesas de castanha possuem qualidades reconhecidas no mercado internacional e dada a sua importância, torna-se cada vez mais necessário programas que monitorizem a sua biodiversidade tendo em vista a preservação do património genético nacional.
A caracterização das proteínas de reserva da castanha é uma importante ferramenta na avaliação da sua variabilidade genética. Neste sentido, propusemos avaliar a aplicabilidade das proteínas de reserva da castanha como marcadores de diversidade genética em Castanea sativa Mill.. Foi realizada electroforese monodimensional (SDS-PAGE) de 21 cultivares portuguesas para definir os seus perfis proteicos, que se revelaram altamente polimórficos para as albuminas e globulinas, tendo sido identificadas 20 e 25 bandas, respectivamente. Este trabalho demonstrou níveis elevados de diversidade inter-varietal e intra-varietal, com valores de similaridade genética a variar entre 0,55 e 0,95.
Através de uma análise ao conteúdo de proteína bruta na semente da castanha, pudemos apurar um valor médio de 7,7g por 100g de matéria seca, para as cultivares em estudo. Também através de uma análise dos aminoácidos pudemos verificar que o aminoácido predominante é o ácido aspártico.
Numa fase posterior do trabalho, foi realizada electroforese bidimensional (2-DE), que permitiu a separação dos péptidos, primeiro pelo seu pI e posteriormente de acordo com sua massa molecular relativa por meio de SDS-PAGE, de modo a obter um proteoma completo das cultivares ‘Judia’ e ‘Rebolão’. Posteriormente através de uma análise por espectrometria de massa, ionização/dessorção a laser assistida por matriz (MALDI-TOF-MS), e bioinformática, foi possível identificar as sequências de proteína presentes e inferir da sua função biológica. Através dos géis 2-DE foi possível detectar 81 spots para a cultivar ‘Judia’ e 174 para a cultivar ‘Rebolão’. A maioria das proteínas identificadas foram relacionadas com funções biológicas, como armazenamento de substratos nutritivos, tolerância ao stress térmico, fixação de carbono, metabolismo de hidratos de carbono, catabolismo da quitina e associados a actividade antifúngica.
de diferentes cultivares e a espectrometria de massa revelou-se crucial na identificação de algumas proteínas presentes na castanha podendo inferir-se acerca das suas funções.
Palavras-Chave: Castanea sativa Mill.; Proteínas de reserva; SDS-PAGE; 2-DE;
The European chestnut (Castanea sativa Mill.) is an agro-forestry specie with great economic importance for Central and Northern regions of Portugal. For many centuries chestnuts provided the staple food in rural populations, because of their good adaptation to soil-climatic conditions, until the appearance of maize and potato.
Portuguese cultivars present several characteristics recognized in the international market, thus it is important to create programs that are capable of monitor its biodiversity considering also the preservation of the national genetic heritage.
Storage proteins characterization in chestnut is an important tool in the evaluation of genetic variability. In this sense, we proposed to evaluate the applicability of storage protein of chestnut as markers of genetic diversity in Castanea sativa Mill. It was performed one-dimensional electrophoresis (SDS-PAGE) of 21 Portuguese cultivars to define their protein profiles, which revealed to be highly polymorphic for albumins and globulins,with 20 and 25 polymorphic respectively. This study demonstrated high levels of inter-varietal and intra-varietal diversity, with genetic similarity values ranging from 0.55 to 0.95.
Through an analysis of crude protein content in the nut, we can calculate an average value of 7.7 g per 100 g of raw chestnuts. Also using an amino acid analysis it can be seen that the predominant amino acid is aspartic acid.
At the second stage, it was performed two-dimensional gel electrophoresis (2-DE) enabling the separation of protein, firstly by their pI and then further separated by molecular weight through SDS-PAGE, in order to obtain the complete proteome of the cultivar Judia and ‘Rebolão’. Subsequent analysis by matrix-assisted laser desorption/ionization mass spectrometry (MALDI-TOF-MS) and bioinformatics it was possible to identify protein sequences present in chestnut and infer their function.
Overall, 71 reproducible protein spots were detected by two-dimensional electrophoresis on ‘Judia’ and 171 protein spots were detected on ‘Rebolão’. The majority of the proteins identified were related to nutrient reservoir activity, chitin catabolic process, carbohydrate metabolic process, fixation of carbon, heat stress tolerance and antifungal activity. The storage proteins analysis in castanea sativa is a useful tool by providing additional information to explain the genetic and adaptive diversity within different cultivars and mass spectrometry proved crucial for the identification of some proteins present in chestnuts.
Orientação ... v
Agradecimentos ... vii
Resumo ... ix
Abstract ... xi
Índice de Figuras ... xv
Índice de Tabelas ... xvii
Abreviaturas ... xix
1. Revisão da literatura ... 1
1.1. A castanha ... 1
1.2. Os Recursos Genéticos Portugueses ... 8
1.3. Monitorização da diversidade genética através de marcadores proteicos ... 12
1.4. Proteómica ... 15
1.5. Ferramentas Proteómicas ... 16
1.5.1. Técnicas de separação proteica ... 16
1.5.2. Espectrometria de massa ... 18
1.5.2.1. Identificação das proteínas utilizando MALDI-TOF/MS ... 19
2. Objectivos ... 21
3. Material e Métodos ... 22
3.1. Descrição do material vegetal ... 22
3.2. Análise monodimensional ... 23
3.2.1. Extracção fraccionada das proteínas de reserva da castanha ... 23
3.2.2. Electroforese monodimensional do tipo SDS-PAGE ... 25
3.2.3. Coloração com Azul de Coomassie R250 ... 27
3.2.4. Análise estatística ... 27
3.3. Análise de aminoácidos ... 28
3.4. Análise bidimensional ... 29
3.4.1. Extracção de proteínas totais de castanha para IEF x SDS-PAGE ... 29
3.4.2. Quantificação proteica ... 30
3.4.3. Electroforese bidimensional do tipo IEF x SDS-PAGE ... 31
3.4.3.1. Rehidratação tiras de gel IPG ... 32
3.4.3.2. Focalização com IPGphor ... 32
3.4.3.3. Equilibração das tiras de gel IPG ... 33
3.4.3.4. Gel 2-DE... 34
3.5. Digestão proteica ... 36
3.6. Análise por MALDI-TOF/MS ... 37
4. Resultados e Discussão ... 38
4.1. Análise monodimensional ... 38
4.2. Análise de aminoácidos ... 45
4.3. Análise bidimensional das cultivares ‘Judia’ 76 e ‘Rebolão’... 48
5. Conclusão ... 59
6. Referências bibliográficas ... 61
Figura 1 - Distribuição geográfica das espécies do género Castanea……….……2 Figura 2 – Os dois eventos migratórios do género Castanea com base na análise filogenética para a
região trnL-trnF de DNA cloroplastidial (Adaptado de Abreu, 2007 [7])………..…..…3
Figura 3 – Distribuição geográfica da produção de castanha no ano 2014 (Adaptado de FAOSTAT,
2017 [12])………...………….4
Figura 4 – Distribuição geográfica das cultivares de castanhas utilizadas neste trabalho…………...22 Figura 5 – Extracção sequencial das proteínas de reserva da castanha (Adaptado de Collada e
colaboradores, 1986 [43])………...24
Figura 6 - Placas e elementos do sistema da tina de electroforese vertical. A) Tampa da tina; B) Tina
de electroforese; C) Bandeja superior; D) Nível; E) Pente; F) Parafusos; G) Espaçador; H) Braçadeira; I) Vidros; J) Suporte; K) Dispositivo de refrigeração. (Fonte: Igrejas, 1996 [69])……….25
Figura 7 – Metodologia de extracção total de proteínas de sementes (Adaptado de Zienkiewicz e
colaboradores, 2014 [74])………...30
Figura 8 – Placa de aço inoxidável onde foram depositadas as amostras e cobertas com a
matriz……….….37
Figura 9 – Perfil electroforético monodimensional da cultivar ‘Longal’ mostrando as diversas fracções
proteicas (albuminas, globulinas, prolaminas e glutelinas………..….38
Figura 10 – Perfil electroforético para as albuminas. M) Marcador de massa molecular relativa;
1-‘Aveleira’, 2-‘Bária’, 3-‘Bebim’, 4-‘Benfeita’, 5-‘Boaventura’, 6-‘Côta’, 7-‘Demanda’, 8-‘Judia’ 144, 9-‘Judia’ 76, 10-‘Lada’, 11-‘Lamela’, 12-‘Longal’, 13-‘Martaínha’ 23, 14-‘Híbrido’, 15-‘Martaínha’ 86, 16-‘Misericórdia’, 17-‘Moreira’, 18-‘Negra’ 136, 19-‘Negral’ 16, 20-‘Passã’, 21-‘Rebolão’, 22-‘Sousã’ 149, 23- ‘Sousã’ 8, 24-‘Trigueira’, 25-‘Verdeal’ 133 e 26-‘Verdeal’ 81………..39
Figura 11 – Perfil electroforético para as globulinas. M) Marcador de massa molecular relativa;
1-‘Aveleira’, 2-‘Bária’, 3-‘Bebim’, 4-‘Benfeita’, 5-‘Boaventura’, 6-‘Côta’, 7-‘Demanda’, 8-‘Judia’ 144, 9-‘Judia’ 76, 10-‘Lada’, 11-‘Lamela’, 12-‘Longal’, 13-‘Martaínha’ 23, 14-‘Híbrido’, 15-‘Martaínha’ 86, 16-‘Misericórdia’, 17-‘Moreira’, 18-‘Negr’a 136, 19-‘Negral’ 16, 20-‘Passã’, 21-‘Rebolão’, 22-‘Sousã’ 149, 23- ‘Sousã’ 8, 24-‘Trigueira’, 25-‘Verdeal’ 133 e 26-‘Verdeal’ 81.………..40
Figura 12 – Diagrama das bandas obtidos pela análise do software CLIQS 1D PRO (Totallab). Em cima
observa-se a sobreposição de todas as bandas para a análise das albuminas e em baixo para as globulinas………..…..40
Figura 13 – Dendrograma representativo da similaridade genética para as albuminas existentes no
conjunto analisado de cultivares de Castanea sativa……….………....….….42
Figura 14 – Dendrograma representativo da similaridade genética para as globulinas existentes no
Figura 16 – Imagem do gel bidimensional da amostra da cultivar ‘Judia’ 76 com coloração Coomassie
G-250. Tira de gel IPG pH 3-10 de 13 cm………..………49
Figura 17 – Imagem do gel bidimensional da amostra da cultivar ‘Rebolão’ com coloração Coomassie
G-250. Tira de gel IPG pH 3-10 de 13 cm………..49
Figura 18 – Géis bidimensionais de referência com marcação de spots obtidos através do software
SameSpots. A - Gel correspondente à cultivar ‘Judia’ 76; B - Gel correspondente à cultivar ‘Rebolão’………50
Figura 29 – Géis de referência com marcação de spots onde se obteve uma identificação proteica. Do
lado esquerdo o gel referente à cultivar Judia e do lado direito o corresponde à cultivar ‘Rebolão’. Os
spots indicados com a seta vermelha apresentaram os péptidos onde ocorreu identificação após a
realização de BLAST……….………….…56
Figura 20 – BLAST da sequência 9577 from Patent WO2010046221 obtida por MALDI-TOF/MS
Tabela 1 – Enquadramento taxonómico do Castanheiro [2]………1
Tabela 2 – Produção de castanha na Europa no ano 2014 (Adaptado de FAOSTAT, 2017 [12]……....5
Tabela 3 – Exportação mundial de castanha no ano 2013 referente aos maiores produtores a nível Mundial. A cinzento encontram-se destacados os países Europeus (Adaptado de FAOSTAT, 2017 [12])………..6
Tabela 4 – Composição centesimal de macronutrientes, fibras e valor energético da castanha portuguesa (g/100g) (Adaptado de Souza e colaboradores, 2014 [19])………...7
Tabela 5 – Distribuição geográfica das diferentes cultivares Portuguesas de castanha (Adaptado de Ferreira-Cardoso e Pimentel-Pereira, 2007 [8])……….……….11
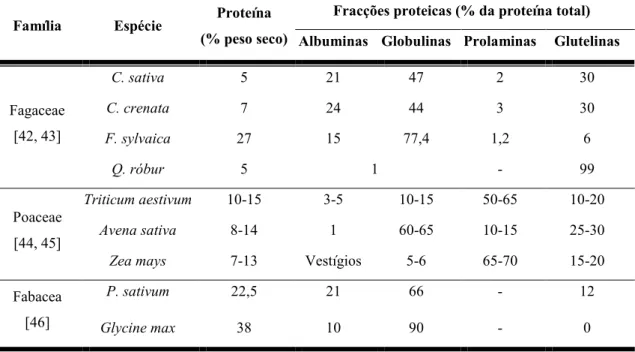
Tabela 6 – Conteúdo proteico, em percentagem do peso total e de diferentes fracções proteicas, das sementes de espécies das famílias Fagaceae, Fabaceae e Poaceae……….15
Tabela 7 – Lista de cultivares de castanhas utilizadas neste trabalho e respectivas origens………….22
Tabela 8 – Preparação dos géis de separação (T: 12,52% e C: 0,97)………..……..26
Tabela 9 – Gel de concentração (T: 2,88% e C: 1,42%)………27
Tabela 10 – Reagentes utilizados no protocolo de quantificação proteica………30
Tabela 11 – Preparação da curva padrão………...………31
Tabela 12 – Parâmetros específicos da focalização isoeléctrica………...…...…………..33
Tabela 13 – Teor de proteína bruta em g por 100 g de matéria seca das cultivares portuguesas de castanha em estudo………...………..….46
Tabela 14 – Média do conteúdo de aminoácidos (g/gN) em matéria seca de castanha……….47
Tabela 15 – Proteínas em que se obteve uma identificação, por MALDI-TOF/MS, imediata de proteínas para a cultivar ‘Judia’ 76………...………..52
Tabela 16 – Proteínas em que se obteve uma identificação, por MALDI-TOF/MS, imediata de proteínas para a cultivar ‘Rebolão’……….53
1-DE - Eletroforese monodimensional, Monodimensional Electrophoresis. 2-DE – Electroforese bidimensional, Two Dimensional Electrophoresis. ACN - Acetonitrilo, Acetonitrile.
APS - Persulfato de amónio, Ammonium persulfate. BSA – Albumina de Soro bovino, Bovine serum albumin.
CHAPS – 3-((3-colamidopropil) dimetilamónio)-1-propanossulfonato. DNA – Ácido desoxirribonucleico, Deoxyribonucleic Acid.
DOP – Denominação de Origem Protegida. DTT – Ditiotreitol, Dithiothreitol.
ESI - Ionização por electrospray, Electrospray ionization.
FAO – Organização das Nações Unidas para a Agricultura e Alimentação, Food and
Agriculture Organization.
IEF – Focalização isoeléctrica, Isoelectric Focusing.
IEF x SDS-PAGE – Focalização isoeléctrica seguida de electroforese em gel de poliacrilamida
na presença do dodecil sulfato de sódio, Isoelectric Focusing followed by Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis.
IPG - Gradiente de pH imobilizado, Immobilized pH gradient. m/z – Relação entre Massa e Carga.
MALDI - Ionização/Dessorção de matriz assistida por laser, Matrix Assisted Laser Desorption
Ionization.
MALDI-TOF - Ionização/Dessorção de Matriz Assistida por Laser – tempo de voo, Matrix
Assisted Laser Desorption Ionization - Time Of Flight.
MALDI-TOF-MS – Ionização/Dessorção de matriz assistida por laser - tempo de voo por
espectrometria de massa, Matrix Assisted Laser Desorption Ionization - Time Of Flight-Mass Spectrometry.
Mr- Massa molecular relativa, Molecular weight.
MS - Espetrometria de massa, Mass spectrometry. NCBI – National Center for Biotechnology Information.
PCR – Reacção em Cadeia por acção da Polimerase, Polymerase Chain Reaction. pI – Ponto isoeléctrico, Isoelectric point.
rpm – Rotações por minuto.
TCA - Ácido tricloroacético, Trichloroacetic acid.
TEMED - Tetrametiletilenediamina, Tetramethylethylenediamine. TFA – Ácido trifluoroacético, Trifluoroacetic acid.
TOF - Tempo de voo, Time of flight.
Tris - Hidroximetil aminometano, Hydroxymethyl aminomethane. Tris-HCl - Tris(hidroximetil)aminometano - Ácido clorídrico. UV - Ultravioleta.
v/v – Razão volume/volume. p/v – Razão peso/volume.
1. Revisão da literatura
1.1. A castanha
O termo Castanea teve origem na Ásia Menor onde é conhecida por Kashtah e provavelmente foi introduzida na Europa através da Grécia, onde a palavra latina Castanea deriva do grego Kastanon. O castanheiro pertence à família das Fagáceas e o seu género é Castanea que engloba cerca de 13 espécies, sendo todas diplóides (X=12; 2n=24) [1].
Segundo Cronquist [2], o castanheiro apresenta o seguinte enquadramento taxonómico (Tabela 1):
Tabela 1 - Enquadramento taxonómico do Castanheiro [2].
Reino: Plantae
Sub-Reino: Embryophyta (Cornophyta)
Divisão: Spermatophyta (Anthophyta)
Sub-Divisão: Angiospermae (Magnoliophytina) Classe: Dicotyledonae (Magnoliopsida) Sub-Classe: Hamamelidae
Ordem: Fagales
Família: Fagaceae Sub-Família: Castaneoideae Género: Castanea
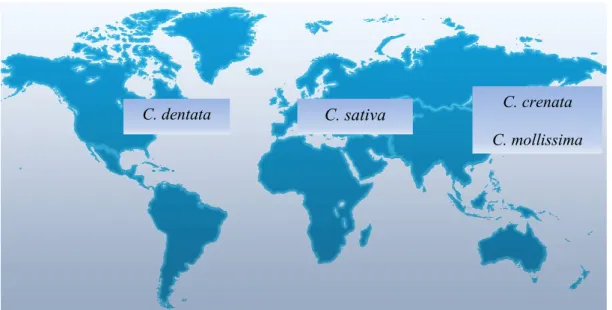
Das cerca de 13 espécies conhecidas apenas 4 têm interesse comercial, encontrando-se distribuídas geograficamente em três principais áreas (Figura 1): Ásia com Castanea crenata Sieb. e Zucc. (Japão) e Castanea mollissima Blume. (China e Coreia), América do Norte com Castanea dentata Borkh. e Europa com Castanea sativa Mill. [3, 4].
Figura 1 – Distribuição geográfica das espécies do género Castanea.
Estas espécies apresentam uma enorme biodiversidade, o que se reflecte na sua adaptação a diferentes condições ambientais. Isto comprova-se pela variabilidade das características morfológicas e ecológicas, ciclos vegetativos e reprodutivos, tamanho do fruto, características da madeira, adaptabilidade e resistência a stresses bióticos e abióticos, onde a conjugação entre biodiversidade natural e selecção humana é muito fraca e por vezes pouco clara [5].
Segundo Paffetti e seus colaboradores [6], admite-se que a China foi o provável centro de dispersão geográfica do castanheiro e ocorreram dois subsequentes eventos de migração intercontinental: da Ásia para a América do Norte e do Médio Oriente para Europa. Além disso, estes autores reportaram através de valores de divergência para a região trnL-trnF de DNA cloroplastidial, que a espécie C. dentata resulta próxima de C. mollissima, enquanto C. crenata se situa entre C. sativa e C. dentata-C. molíssima. Daí, a maior divergência entre C. dentata e C. sativa (Figura 2).
C. dentata C. sativa C. crenata
Figura 2 – Os dois eventos migratórios do género Castanea com base na análise filogenética para
a região trnL-trnF de DNA cloroplastidial (Adaptado de Abreu, 2007 [7]).
Em Portugal, como no resto da Europa, Castanea sativa Mill., também conhecido por Castanheiro Europeu, é a espécie mais cultivada e engloba inúmeras cultivares produtoras de fruto com considerável valor comercial [8]. Além disso, também é cultivada Castanea crenata (Sieb e Zucc) como porta-enxerto de Castanea sativa por apresentar vantagens no desenvolvimento e na resistência a doenças [9].
Castanea sativa é uma espécie de folha caduca que atinge grandes dimensões, cerca de 20-30 metros de altura, de copa semi-esférica, ligeiramente alongada, sendo uma das árvores mais imponentes dos nossos ecossistemas florestais. Apresenta um tronco espesso, liso nos primeiros 10-15 anos, mas rapidamente a casca se fendilha criando linhas pouco profundas, que com o envelhecimento faz com que o tronco se torne torcido. Com folhas verdes brilhantes de grande tamanho, lanceoladas e dentadas, dispostas alternadamente sobre os ramos. A sua semente, conhecida por castanha, surge no interior do ouriço e o seu conjunto constitui o fruto do castanheiro [9, 10].
Durante muitos séculos, no Norte de Portugal, a castanha teve um importante papel económico e social. Constituiu a base alimentar das populações rurais, devido à sua boa adaptação às condições edafoclimáticas, até ao aparecimento do milho e da batata.
O aparecimento de doenças como a “tinta do castanheiro” (Phytophthora) e o “cancro do castanheiro” (Cryphonectria parasitica) também contribuiu de forma drástica para o declínio da sua produção, assim como o aumento de incêndios que devastou muitos hectares [11].
O castanheiro é também uma excelente fonte de madeira, de elevada qualidade, sendo utilizada em mobiliário, soalhos, construção naval, decoração de interiores, etc. Quando o objectivo das plantações é a produção de madeira são utilizados castanheiros bravos, designando-se o povoamento por castinçal; quando o objectivo é a produção frutícola são cultivados castanheiros mansos e atribui-se o nome de souto [1].
Actualmente, a castanha tem vindo a recuperar um lugar de destaque na gastronomia, devido ao reconhecimento do seu valor nutricional, tornando-se muito procurada pelo consumidor [1].
Segundo os dados recentes obtidos pela Organização das Nações Unidas para Alimentação e Agricultura (FAO, Food and Agriculture Organization of the United Nations) referentes a 2014, a China apresenta-se como o maior produtor Mundial de castanha com enorme destaque. Portugal apresenta-se em oitavo lugar, sendo um dos principais países produtores a nível Mundial [12] (Figura 3).
Figura 3 – Distribuição geográfica da produção de castanha no ano 2014 (Adaptado de
FAOSTAT, 2017 [12]). 1.684 28 57 16 64 78 52 21 12 9 Portugal Espanha França Grécia Turquia China Bolívia Japão Coreia do Norte Coreia do Sul Itália
Produção >1.000 milhares de toneladas Produção entre 50-100 milhares de toneladas Produção entre 20-50 milhares de toneladas Produção até 20 milhares de toneladas
Contudo, se olharmos apenas para os maiores produtores Europeus no ano de 2014, que predominantemente cultivam C. sativa Mill., Portugal situa-se em terceiro lugar com cerca de 18.465 toneladas produzidas (Tabela 2). A Itália encontra-se como maior produtor Europeu, produzindo mais do dobro de Portugal. Já em segundo lugar encontra-se a Grécia, com uma produção superior à de Portugal. A Espanha com uma produção inferior à de Portugal, engloba assim os quatro maiores produtores europeus. Com uma menor produção encontra-se a Albânia e França, que apesar de apresentarem uma produção bastante inferior à de Portugal já é bastante significativa no seio da Europa (Tabela 2)
Tabela 2 – Produção de castanha na Europa no ano 2014 (Adaptado de FAOSTAT, 2017 [12]).
País Produção em toneladas
Itália 51.959 Grécia 28.440 Portugal 18.465 Espanha 16.136 França 8.668 Albânia 6.590
Ao nível das exportações de castanha, com base em dados da FAO referentes ao ano de 2013, a China foi o país que mais exportou, estando Portugal em segundo lugar com cerca de 16 mil toneladas, representando 65,4% da produção. A nível Europeu, Itália e Espanha também se destacaram e é de salientar que Espanha exportou quase tanto quanto produziu.
Apesar de a China dominar as exportações, estas apenas representam cerca 2,3% da sua produção e o valor comercial pago por tonelada é quase cerca de metade relativamente aos países Europeus. Já Portugal exportou cerca de 65,3% da sua produção total, contribuindo com cerca de 62 milhões de dólares (Tabela 3).
Tabela 3 - Exportação mundial de castanha no ano 2013 referente aos maiores produtores a nível
Mundial. A cinzento encontram-se destacados os países Europeus (Adaptado de FAOSTAT, 2017 [12]).
País Produção (t) Exportação (t) Exportação (1000 USD) Valor (USD)/kg Exportada
China 1719410 39120 84912 2,17
Coreia do Sul 64184 12285 30369 2,47
Turquia 60019 5166 18449 3,57
Bolívia 76035 N/A N/A N/A
Itália 55086 14148 80784 5,71 Grécia 27800 1650 6201 3,76 Portugal 24739 16153 61960 3,84 Japão 21000 1314 3625 2,76 Espanha 17200 13690 42980 3,14 França 9200 3716 21407 5,76
A área de cultivo de castanha em Portugal corresponde a cerca de 35 mil hectares e prevê-se o aumento nos próximos anos devido ao incentivo da sua produção e ao fácil escoamento que este produto apresenta nos mercados [12]. Além disso, algumas empresas portuguesas especializadas na transformação e comercialização de castanha têm sido fundamentais para o escoamento da castanha nacional para o mercado internacional [13]. No que diz respeito à comercialização da castanha em Portugal verifica-se três tipos de mercados: o mercado de frutos secos, exportação em fresco ou congelada e o mercado de transformação. Existem mais de 300 produtos derivados da castanha, desde purés, compotas, iogurtes, cerveja, farinha, etc.
As qualidades nutricionais quer da castanha quer da farinha de castanha revelam-se muito interessantes. Aprerevelam-sentam uma importante riqueza em hidratos de carbono: amido, composto por 33% de amilose e 67% de amilopectina [14]. Constituem mais de metade da matéria seca total, traduzindo-se num importante valor calórico, o que faz da castanha uma excelente fonte energética, ideal para prevenir situações de desgaste físico e intelectual. As duas formas de amido apresentam efeitos positivos para a saúde, como fornecimento de energia (catabolismo da amilose e amilopectina em glicose) e também funções a nível gástrico devido ao catabolismo bacteriano de amilopectina [14].
A castanha fresca, quando comparado com outras sementes comestíveis, apresenta valores de teor de proteína bruta relativamente baixos. No entanto, estes dados
revelam-se importantes biologicamente, pois possuem quarevelam-se todos os aminoácidos esrevelam-senciais (Tabela 4) [14].
A castanha apresenta também baixo teor de gordura quando comparada a outras sementes comestíveis, sendo pobre em ácidos gordos saturados, isenta de colesterol e parcialmente rica em ácidos gordos insaturados (Tabela 4) [14, 15].
Relativamente à fibra alimentar, a castanha apresenta concentrações favoráveis comparando com outras sementes. As fibras são responsáveis pela manutenção da actividade intestinal e da sua microflora (Tabela 4) [14].
Esta semente possui na sua constituição várias vitaminas, destacando-se a vitamina E e vitamina C. A vitamina E desempenha funções antioxidantes no organismo. Já a vitamina C participa em processos celulares de oxidação redução e é importante na biossíntese de catecolaminas [16]. Em menores quantidades também são encontradas vitaminas como: tiamina, riboflavina, niacina, piridoxina e folato [14].
Este alimento é uma boa fonte de minerais, como o cobre, ferro, manganês, magnésio, potássio e selénio [17]. Destacando-se a quantidade de ferro, que segundo Borges e seus colaboradores [18] a média presente em 100g de castanha é cerca de 8,1 mg, representando 100% da dose diária recomendada para os homens e 44% para as mulheres.
Tabela 4 - Composição centesimal de macronutrientes, fibras e valor energético da castanha
portuguesa (g/100g) (Adaptado de Souza e colaboradores, 2014 [19]).
Castanha/semente comestível
Componentes g/100g kcal
Hidratos de
Carbono Proteínas Lípidos Alimentar Fibra Energético Valor
Castanha Portuguesa 41,5 6,15 2,4 22,85 191
Amendoim 12,01 24,03 44,57 11,30 545,29
Avelã 2,57 14,77 63,18 12,88 637,98
Amêndoa de baru 10,95 26,44 41,04 13,90 518,04
Castanha de caju 32,08 18,81 42,06 N/A 582,10
Castanha do Pará 6,27 14,11 64,94 8,02 665,98
Pistache 25,42 19,80 45,83 N/A 593,35
Macadâmia 22,18 13,81 66,16 N/A 717,76
Este conjunto de dados demonstram a importância do valor nutricional da castanha e da sua relevância na economia nacional ao nível das exportações, que devido
à excelente qualidade das variedades de castanha tornam o produto português muito procurado a nível internacional, tanto para a sua industrialização como para o consumo em fresco.
1.2. Os Recursos Genéticos Portugueses
Dada a importância da castanha, foram criadas Denominações de Origem Protegida (DOP), em que o principal objectivo é a preservação e valorização do património genético do castanheiro, fixando regras de produção que mantenham os parâmetros diferenciadores da produção, nas diferentes regiões, e regras de mercado, enquadrando os produtos regionais de qualidade como factores de desenvolvimento local [11]. A nível nacional existem quatro “Denominações de Origem Protegida” (DOP) [20]:
‘Castanha da Terra Fria’- são considerados os frutos das cultivares ‘Longal’, ‘Judia’, ‘Côta’, ‘Amarelal’, ‘Lamela’, ‘Aveleira’, ‘Boa Ventura’, ‘Trigueira’, ‘Martaínha’ e ‘Negral’. Mais de 70% da produção corresponde à cultivar ‘Longal’, sendo os restantes 30% relativos à produção das outras cultivares mencionadas. A sua área de produção está limitada aos concelhos dos distritos de Bragança e Vila Real.
‘Castanha dos Soutos da Lapa’- são utilizadas as cultivares ‘Martaínha’ e ‘Longal’. A área geográfica de produção abrange alguns concelhos dos distritos de Viseu e da Guarda. Cerca de 58% da área plantada de castanheiro pertence aos concelhos de Trancoso e Penedono, e mais de 90% da produção deverá corresponder à cultivar ‘Martaínha’.
‘Castanha da Padrela’- castanhas obtidas a partir das cultivares ‘Judia’, ‘Longal’, ‘Lada’, ‘Negral’, ‘Côta’ e ‘Preta’. A área geográfica abrange algumas freguesias dos concelhos de Chaves, Murça, Vila Pouca de Aguiar e de Valpaços, correspondendo este último a cerca de 80% da área plantada. Aproximadamente 80% da produção corresponde à cultivar ‘Judia’ e 20% da produção às restantes cultivares.
‘Castanha de Marvão’- tem origem nas cultivares ‘Bária’, ‘Colarinha’ e ‘Bravo’. A área geográfica abrange os concelhos de Marvão, Castelo de Vide e Portalegre.
Em Portugal existe um número extenso de cultivares regionais de castanheiros que possui uma grande variabilidade genética, sendo algumas mais estáveis do que outras. As cultivares Portuguesas são policlonais, ou seja, uma mesma cultivar apresenta mais do que um genótipo, o que indica que se propagou mais que um clone da mesma cultivar [11].
Estudos genotípicos demonstram diferentes genótipos para diferentes cultivares, que resultam de mutações e polinizações cruzadas. Verifica-se que existem cultivares com genótipos muito repetidos que consequentemente poderão considerar-se como o clone principal, a partir dos quais derivam outros clones dentro da mesma cultivar ou até originam outras cultivares, por semente.
Estudos de polinização no castanheiro revelam que algumas características biométricas e fisiológicas, podem ser afectadas pelo genótipo do polén da cultivar polinizadora, isto é, pelas características do progenitor masculino, traduzindo assim fenómenos de xenia e metaxenia [21, 22].
Todavia, Gomes e colaboradores [22] referem que o factor mais determinante entre o tamanho e forma do fruto, considerando o mesmo local e genótipo, é a pluviosidade e o modo como ela se distribui, principalmente, na época mais seca do ano, que coincide com o período de maior actividade vegetativa desta espécie.
Inicialmente a nomenclatura das cultivares de castanha foi atribuída tendo por base o seu fenótipo, ou seja, com base na observação das suas características morfológicas. Nos últimos anos, devido a realização de estudos genotípicos demonstrou-se existirem relações específicas entre cultivares regionais e nacionais, gerando uma certa confusão quanto às cultivares de castanheiro. Apesar da existência de muitas cultivares de castanha em Portugal, têm sido atribuídas designações diferentes para a mesma cultivar com localização geográfica próxima. Outros problemas surgiram quando algumas cultivares possuíam origens e designações locais aparentemente diferentes, mas na realidade derivavam da mesma cultivar. Este tipo de confusão ocorre quando as cultivares divergem morfologicamente devido a factores ambientais [23].
Vários estudos têm sido realizados para combater estes problemas de nomenclatura das cultivares Portuguesas de castanha. Por exemplo, Dinis e colaboradores [24] realizaram um estudo com o objectivo de avaliar a heterogeneidade do genótipo da cultivar ‘Judia’ em Trás-os-Montes, recorrendo ao uso de marcadores de sequências simples repetidas (SSR, Single Sequence Repeats) para 25 amostras de localizações distintas, onde encontraram algumas variações intra-varietais.
Noutro estudo realizado por Pimentel-Pereira e colaboradores [26], a nível isoenzimático, foram usados 7 tipos de marcadores, que permitiram a discriminação de 32 classes genotípicas. Cada classe correspondeu a uma ou mais cultivares, sendo em alguns casos geneticamente muito semelhantes e com designações diferentes (sinonímia), e noutros casos eram cultivares geneticamente distintas, com a mesma designação e correspondiam a classes diferentes (homonímia).
Trabalhos de caracterização molecular de cultivares de castanha portuguesa realizados por Costa e colaboradores [25] e Ribeiro [26], apresentam um possível caso de heteronímia entre as cultivares ‘Martaínha’ e ‘Verdeal’, justificada pelo valor de similaridade de 82% entre as duas cultivares. Estes trabalhos referem ainda a instabilidade do ponto de vista genotípico das cultivares ‘Longal’, ‘Martaínha’, ou seja, apresentam grande variabilidade intra-populacional (inter-varietal).
Pimentel-Pereira e colaboradores [27] destacam os seguintes grupos de cultivares, que não poderão ser mais que designações diferentes para um mesmo genótipo:
-‘Riscal’ e ‘Verdeal’ (provenientes de Armamar); -‘Cancela’ (Lamego) e ‘Negral’ (Valpaços);
-‘Lamela’ (Vinhais) e ‘Pelada’ (Moimenta da Beira); -‘Cota’ (Murça) e ‘Negral’ (Moimenta da Beira); -‘Cota’ (Valpaços) e ‘Demanda’ (Tarouca); -‘Pelada’ (Boticas) e ‘Redonda’ (Montalegre).
Com base numa compilação de trabalhos realizados de detecção e inventariação das diferentes cultivares Portuguesas de castanha foi possível detalhar a distribuição das mesmas, como podemos observar na Tabela 5 [8].
Tabela 5 - Distribuição geográfica das diferentes cultivares Portuguesas de castanha (Adaptado
de Ferreira-Cardoso e Pimentel-Pereira, 2007 [8]).
Distrito Concelho Cultivares
Braga Terras de Bouro ‘Amarelal’; ‘Misericórdia’
Bragança
Alfândega da Fé ‘Enxerta’
Bragança ‘Enxerta’; ‘Judia’ Carrazeda de Ansiães ‘Longal’; ‘Enxerta’ Macedo de Cavaleiros ‘Enxerta’
Mogadouro ‘Enxerta’; ‘Negra’ Vinhais
‘Judia’ (Carrazeda); ‘Aveleira’; ‘Lamela’; ‘Trigueira’; ‘Boaventura’; ‘Longal’
Guarda Trancoso ‘Martaínha’; ‘Longal’
Portalegre Marvão ‘Bária’; ‘Colarinha’; ‘Enxerta’ (Longal)
Vila Real
Boticas ‘Pelada’; ‘Galizã’; ‘Longal’
Chaves ‘Vermelha’; ‘Longa’
Montalegre ‘Redonda’; ‘Nacional’; ‘Enxerta’; ‘Longal’
Murça ‘Côta’; ‘Longal’
Valpaços
‘Judia’; ‘Lada’; ‘Negral’; ‘Sousão’; ‘Lamela’; ‘Rebolão’; ‘Côta’;
‘Soutinha’; ‘Longal’ Vila Pouca ‘Côta’; ‘Longal’; ‘Bairral’ Vila Real (Campeã) ‘Benfeita’; ‘Bebim’; ‘Moreira’;
‘Longal’
Viseu
Armamar ‘Riscal’, ‘Longal’; ‘Serrano’ Lamego ‘Cancela’; ‘Longal’; ‘Rabodiça’
Meda ‘Verdeal’; ‘Longal’
Moimenta da Beira ‘Negra’; ‘Demanda’; ‘Carreiró’; ‘Longal’
Penedono ‘Passã’; ‘Martaínha’; ‘Longal’
Relativamente ao valor frutícola e comercial, as cultivares portuguesas podem organizar-se da seguinte forma [8]:
‘Judia’, ‘Longal’, ‘Lamela’, ‘Aveleira’ e ‘Boaventura’, no distrito de Bragança; ‘Judia’, ‘Longal’, ‘Lada’, ‘Negral’, ’Cota’ e ‘Benfeita’, no distrito de Vila Real; ‘Martaínha’ e ‘Longal’ nos distritos de Viseu e Guarda;
‘Bária’, ‘Colarinha’ e ‘Enxerta’ no distrito de Portalegre.
1.3. Monitorização da diversidade genética através de marcadores proteicos
Os recursos genéticos vegetais representam a base para o desenvolvimento da agricultura e um reservatório da adaptação genética que actua como um tampão contra as alterações ambientais. Eles fornecem material em bruto que, quando usados de maneira correcta, produzem novas e melhores cultivares, e são uma fonte insubstituível de características como resistência a doenças e pragas, adaptação a diferentes ambientes e maiores produtividades [28, 29].
No que respeita ao castanheiro europeu, que se observa como uma espécie largamente difundida e que tem sido cultivada pelo Homem por um longo período de tempo, a sua variabilidade genética tem sido fortemente reduzida nas populações naturais, bem como nos povoamentos cultivados. Tal deve-se às perturbações ambientais, bem como aos fortes ataques dos parasitas a que está sujeito, nas últimas décadas [30].
Neste sentido, acresce a necessidade de programas que façam a monitorização da biodiversidade e projectem a sua preservação e exploração tecnológica e económica, pois impedir a perda da mesma deve ser um objectivo para a sociedade.
Qualquer “sinal”, desde um fenótipo, um fragmento DNA, uma proteína, cuja presença num conjunto de genótipos se manifesta como polimórfica é considerada como marcador genético, sendo susceptível de ser utilizado em combinação com outros marcadores, como característico de um determinado genótipo [31].
Solanille [32] classificou os marcadores genéticos em quatro grupos: morfológicos, bioquímicos, moleculares e citogenéticos. De acordo com Lefebvre e Chèvre [33] marcadores genéticos podem agrupar-se em dois grandes grupos: marcadores morfológicos e marcadores moleculares. Dentro destes últimos podem distinguir-se os marcadores bioquímicos (proteínas enzimáticas ou não enzimáticas) e marcadores de
DNA que podem ser obtidos mediante hibridação do DNA e baseados nas técnicas de reacção em cadeia da polimerase (PCR, Polymerase Chain Reaction).
As proteínas de reserva (proteínas não enzimáticas) têm sido utilizadas como importantes marcadores genéticos em várias espécies, devido ao seu elevado polimorfismo, o controlo genético simples, a sua independência ambiental e uma análise de resultados fácil, rápida e económica [34].
A análise de alelos de proteínas de reserva é bem conhecida por ser uma ferramenta robusta na genotipagem dos recursos genéticos, nomeadamente em cereais, como o trigo [35].
Apesar de outros métodos, tais como a PCR, poderem fornecer ferramentas interessantes em estudos de genotipagem de recursos genéticos, a utilização de electroforese em gel de poliacrilamida na presença de dodecil sulfato de sódio (SDS-PAGE, Sodium Dodecyl Sulfate - Polyacrylamide Gel Electrophoresis) continua a ser um método valioso, eficiente e económico para a identificação de alelos e distinção de genótipos [35].
A SDS-PAGE é um método de separação de proteínas de acordo com a sua massa molecular relativa, onde as amostras proteicas migram em géis com uma matriz de acrilamida pela aplicação de um campo eléctrico. Nesta técnica as amostras são tratadas com SDS, que tem como objectivo a desnaturação das proteínas e indução de carga negativa, pois estas são anfotéricas, ou seja, o seu pH varia consoante o meio onde estão inseridas. Após este processo, é possível visualizar e identificar as proteínas separadas em bandas pela diferença de massa molecular relativa através da coloração dos géis [36]. Assim, de forma a avaliar a diversidade genética do castanheiro europeu, uma possível abordagem passa pela utilização de proteínas como potenciais marcadores de diversidade genética.
As proteínas de sementes foram classificadas, empiricamente, de acordo com a sua solubilidade e extracção numa série de solventes [37]: albuminas, solúveis em água; globulinas, solúveis em soluções salinas (NaCl a 10% frequentemente usado), mas insolúveis em água; prolaminas, solúveis em etanol a 70% e as glutelinas insolúveis em todas as soluções anteriores, mas solúveis em ácido acético a 0,1M.
Este tipo de classificação ainda hoje é mantido apesar de terem sido incorporadas algumas mudanças no procedimento de extracção sequencial, nomeadamente a adição de um agente redutor (2-mercaptoetanol) para a extracção das prolaminas e a adição de um detergente (geralmente o SDS) e um agente redutor para a extracção das glutelinas [38].
Uma classificação mais recente divide as proteínas de sementes em proteínas de reserva, proteínas estruturais e proteínas biologicamente activas [39].
As proteínas de reserva têm como função biológica fornecer um reservatório de nutrientes essenciais. As proteínas de reserva atingem valores elevados do azoto total da semente, onde se encontra a maior acumulação. São estas as proteínas sintetizadas durante a formação da semente e permanecem estáveis desde a maturação até ao momento da germinação, sendo posteriormente degradadas, funcionando como fonte de carbono, enxofre e azoto para o crescimento das plântulas [40, 41].
As proteínas de reserva mais estudadas dizem respeito a espécies de angiospérmicas pertencentes à família Poaceae (gramíneas) e Fabaceae (leguminosas), salientando que o teor total de proteínas nas sementes em matéria seca varia desde 7% a 38%, respectivamente. Já as espécies de Fagaceaes possuem valores que variam entre 5% a 28%, apresentando a espécie em estudo valores de cerca de 5% (Tabela 6).
Dentro da família Fagaceae existem diferenças nas proteínas de reserva para os diferentes géneros, enquanto em F. sylvaica e Castanea as principais proteínas de reserva são as globulinas, em Quercus estas aparecem em valores muito baixos sendo neste caso as glutelinas as proteínas de reserva [42].
No caso da espécie C. sativa as globulinas são as principais proteínas de reserva com valores de cerca 46,9% da proteína total. Comparando C. sativa com C. crenata, as globulinas demonstram grande heterogeneidade. C. sativa apresenta os seus principais componentes com massa molecular relativa de cerca de 25 kDa e pI próximo de 8,5 [43]. As percentagens das fracções proteicas para o total de proteína presentes na semente são cerca de 21,1% para as albuminas, 29,9% para as glutelinas e 2,1% para as prolaminas (Tabela 6).
Tabela 6 – Conteúdo proteico, em percentagem do peso total e de diferentes fracções proteicas,
das sementes de espécies das famílias Fagaceae, Fabaceae e Poaceae.
Família Espécie Proteína
(% peso seco)
Fracções proteicas (% da proteína total) Albuminas Globulinas Prolaminas Glutelinas
Fagaceae [42, 43] C. sativa 5 21 47 2 30 C. crenata 7 24 44 3 30 F. sylvaica 27 15 77,4 1,2 6 Q. róbur 5 1 - 99 Poaceae [44, 45] Triticum aestivum 10-15 3-5 10-15 50-65 10-20 Avena sativa 8-14 1 60-65 10-15 25-30
Zea mays 7-13 Vestígios 5-6 65-70 15-20
Fabacea [46]
P. sativum 22,5 21 66 - 12
Glycine max 38 10 90 - 0
Estudos anteriores foram realizados com sucesso no uso de proteínas de reserva para avaliar a diversidade genética em Castanea sativa Mill., nomeadamente através da análise de albuminas [47] e globulinas [34].
1.4. Proteómica
O dogma central da biologia permitiu a criação de diferentes áreas de estudo, ómicas, que englobam a caracterização colectiva das mesmas, como a genómica, tanscriptómica, proteómica, metabolómica, etc. A proteómica, como o próprio nome indica, refere-se ao estudo das proteínas, que consiste na identificação, expressão e caracterização proteica.
Sendo as proteínas um dos principais produtos dos genes, a proteómica é uma área em forte expansão, uma vez que está relacionada directamente com a genómica e também com a transcriptómica. Assim, através da correlação entre a genómica e a proteómica é possível estudar e relacionar a expressão génica e a sua correspondência ao nível da expressão proteica, tal como os mecanismos de inibição e expressão de uma proteína, bem como os próprios níveis de expressão das proteínas [48]. Por outro lado, devido à existência de modificações pós-traducionais, a proteómica e a transcriptómica podem ser duas áreas divergentes [49]. A proteómica assenta no estudo das proteínas expressas por
um determinado organismo e o seu proteoma depende da sequência, estrutura e localização das proteínas, modificações pós-transcripcionais, interacção entre proteínas e o processo biológico no qual participam [49].
A cooperação de diferentes áreas como a biologia molecular e bioinformática, é essencial para a compreensão dos processos biológicos das proteínas, sendo estas importantes na criação e manutenção de um sistema biológico [50]. Diversas áreas como a sequenciação de genomas, a separação de proteínas, a espectrometria de massa e a bioinformática têm sido os principais pilares de suporte para as tecnologias usadas em proteómica [51].
1.5. Ferramentas Proteómicas
1.5.1. Técnicas de separação proteica
Existe uma variedade de processos de separação de proteínas que têm sido utilizados previamente a uma análise por espectrometria de massa (MS). A separação de proteínas pode ser realizada consoante a sua carga eléctrica, a sua massa molecular relativa ou a sua afinidade com um substrato. A electroforese tem sido dos processos mais utilizados e melhor adaptados, assim como a cromatografia líquida.
A electroforese em gel poliacrilamida na presença de dodecil sulfato de sódio (SDS-PAGE) é uma técnica que se baseia na separação das proteínas através da sua massa molecular relativa. SDS-PAGE é o meio de separação mais utilizado em proteómica, devido à sua elevada capacidade de separar grandes quantidades de proteínas [52]. Nesta metodologia é utilizado o SDS, um detergente, responsável pela quebra de pontes de hidrogénio e eliminação de interacções hidrofóbicas, permitindo a estabilização das proteínas na sua forma desnaturada. Os polipéptidos adquirem carga negativa conferida pelas moléculas de SDS, tornando-se passíveis de migração ao longo de um gel de poliacrilamida quando exercida uma corrente eléctrica. Os péptidos com maior massa molecular ficam retidos mais cedo no gel e os de menor massa ficam depositados no fundo do gel [53]. A grande desvantagem desta técnica é o limite de resolução, visto que o excesso de bandas podem levar à sua sobreposição, dificultando a sua interpretação. A SDS-PAGE é por isso mais eficiente na análise de um reduzido número de proteínas [54]. A SDS-PAGE quando combinada com a focalização isoeléctrica (IEF) representam uma forte ferramenta de análise de misturas de proteínas extraídas de uma
amostra biológica, pois permite a separação das proteínas pelo seu ponto isoeléctrico (1ª dimensão) e permite também a separação pela sua massa molecular relativa (2ª dimensão), electroforese bidimensional (IEF x SDS-PAGE ou 2-DE) [53]. Esta técnica foi descrita inicialmente por O’Farrell [55] e a base do seu elevado poder de resolução são precisamente as duas dimensões, ou seja, as proteínas serem separadas sequencialmente por dois critérios físicos.
A focalização isoeléctrica requer a utilização de anfólitos por ser um método com capacidade de separar as proteínas de acordo com o ponto isoeléctrico. Uma vez que as proteínas são moléculas anfotéricas, estas podem apresentar diferentes cargas dependendo da sua composição em aminoácidos e do pH que as rodeia. Consoante a relação do pH do tampão com o pI da proteína, esta move-se ou permanece na fase estacionária (pH=pI) [53]. O ponto isoeléctrico de uma proteína é o valor específico de pH em que a carga eléctrica líquida da proteína é zero, de modo a estabilizar através da aplicação de um campo eléctrico [56]. A presença de um gradiente de pH é por isso essencial na focalização isoeléctrica, pois vai permitir que as proteínas se movam para a posição do gradiente onde a sua carga é zero, quando aplicado um campo eléctrico. A focalização isoeléctrica é realizada ainda na presença de ureia, detergentes iónicos e ditiotreitol (DTT). A ureia é responsável pela quebra das ligações de hidrogénio, no entanto não afecta a carga eléctrica intrínseca das proteínas.
Tanto para os géis monodimensionais ou bidimensionais existe uma variedade de métodos para a sua coloração, e respectiva marcação das proteínas, sendo o azul de Coomassie e o nitrato de prata os mais utilizados. As vantagens do azul de Comassie em relação ao nitrato de prata são, essencialmente, a sua simplicidade de execução e compatibilidade com os protocolos de espectrometria de massa, enquanto que o nitrato de prata apesar de possuir um maior limite de detecção revela incompatibilidades com alguns protocolos de MS [57]. As limitações desta técnica estão relacionadas com problemas de resolução e de reprodutibilidade na separação de determinadas proteínas, como é o caso de proteínas muito ácidas ou muito básicas, apresentando problemas na solubilização [58].
1.5.2. Espectrometria de massa
Nas últimas décadas, a espectrometria de massa tornou-se uma das técnicas mais preponderantes na detecção e caracterização de proteínas [51]. A espectrometria de massa tem sido considerada um dos pilares da proteómica. Apresenta uma elevada capacidade na identificação proteica com base em fingerprints de massa peptídica e sequências de aminoácidos, apresenta uma elevada sensibilidade na detecção proteica, bem como elevada capacidade de throughput e integração com métodos quantitativos de identificação de proteínas diferencialmente expressas [51].
A espectrometria de massa é composta por três unidades funcionais: uma fonte de iões que ioniza e transfere as moléculas até à fase de gás, um analisador de massa que separa os iões de acordo com uma razão massa/carga (m/z) e um detector que capta e transforma o sinal em corrente eléctrica, onde a extensão do sinal eléctrico em função da razão m/z é convertida por um processador de dados, dando origem a um espectro de massa correspondente [59]. O analisador de massa é ponto fulcral da tecnologia. Os principais parâmetros da espectrometria de massa, no contexto da proteómica, são: a sensibilidade, resolução, precisão de massa e a capacidade em gerar espectros de massa ricos em informação.
Ao longo dos anos foram sendo desenvolvidos diferentes métodos clássicos de ionização, destacando-se a ionização química e o impacto de electrões. A ionização por electrodifusão (ESI) e a ionização/dessorção a laser assistida por matriz (MALDI) são as duas técnicas de ionização mais utilizadas em espectrometria de massa. Ambas as técnicas permitem a passagem das amostras da fase condensada para a fase gasosa por ionização, mas de acordo com princípios diferentes, visto que no ESI os péptidos ionizados são gerados a partir de uma fase líquida, enquanto no MALDI os péptidos ionizados são gerados a partir de uma matriz sólida [59, 60].
Actualmente existem quatros tipos de analisadores de massa usados em estudos proteómicos: ion trap, time-of-flight (TOF), quadropole e Fourieir transform ion cyclotron (FT-MS).
1.5.2.1. Identificação das proteínas utilizando MALDI-TOF/MS
A espectrometria de massa por MALDI é uma ferramenta analítica para péptidos, proteínas e muitas outras biomoléculas, muito difundida. Na análise por MALDI-TOF/MS, inicialmente o analito é co-cristalisado com uma matriz apropriada, com a capacidade de absorver a luz UV do laser. Uma matriz de MALDI tem de obedecer a dois propósitos fundamentais, capacidade de absorver a energia dos fotões procedentes do laser e transferi-la como energia de excitação para o sistema funcionar como solvente da amostra, reduzindo as forças intermoleculares e impedindo a agregação das moléculas do analito [61].
A mistura matriz-analito recebe um feixe de laser pulsado, ionizando indirectamente as moléculas. O feixe de laser foca apenas uma porção da matriz, de modo a prevenir a destruição da amostra e para aumentar a sensibilidade [62]. As moléculas da matriz são excitadas por ressonância, absorvendo essa energia proveniente do laser e transferindo-a para o analito acidificado, através de um processo de protonação (a matriz transfere os protões acídicos). Consequentemente, a excitação electrónica das moléculas da matriz que absorvem a energia do laser, provoca a dessorção da matriz e dos iões [M+H]+ do analito para o estado gasoso, voando até ao analisador por um tubo, em vácuo. Esta ionização necessita de várias centenas de disparos do laser para alcançar uma razão de signal-to-noise plausível [63]. O analisador de massa TOF determina razões de massa por carga, separando os iões de acordo com a diferença de tempo entre o sinal de partida (ponto de formação) e o pulso gerado quando o ião atinge o detector (ponto de detecção), denominando-se tempo de voo (TOF) [64]. A aplicação de um campo electroestático a um material ionizado gera um ião com carga (z) e com aceleração, suportando o tempo de voo neste princípio. Os iões movem-se no tubo de vácuo dependendo exclusivamente da energia cinética do passo de aceleração [62]. O tempo de voo exigido para chegar ao detector é dependente da massa (m) e carga (z) do analito e é proporcional à raiz quadrada m/z. Assim sendo, quanto mais rápido e leve for o ião, mais rápido atinge o detector, e consequentemente, menor é o seu tempo de voo. Os analitos com m/z diferentes que compõem a amostra complexa são separados, de acordo com o seu tempo de voo, criando um espectro de massa caracterizado pelo m/z e a intensidade dos iões. Normalmente, o MALDI produz iões com carga única (z=1), demonstrando que o m/z de um analito corresponde ao valor da sua massa [65].
Como resultado da sua simplicidade, excelente precisão de massa, alta resolução, sensibilidade e tolerância a contaminantes, o MALDI-TOF seve para identificar proteínas, por aquilo que é conhecido como mapeamento de péptidos, também referido como peptide mass fingerprinting. Neste método, as proteínas são identificadas pela correspondência entre uma lista experimental de massas peptídicas e uma lista de todas as massas calculadas de cada péptido através do acesso a uma base de dados específica, ou seja, massas obtidas no passo anterior são comparadas com bases de dados proteicas. É de salientar que esta técnica é relativamente rápida e permite a obtenção de uma grande quantidade de dados em pouco tempo, partindo de uma quantidade reduzida de amostra [66].
2. Objectivos
Neste trabalho pretendeu-se caracterizar algumas das diferentes cultivares portuguesas de castanha através das proteínas presentes na sua semente. Para tal propusemo-nos:
- Analisar o potencial das proteínas de reserva de castanha como uma via para estimar a distância genética entre as cultivares de Castanea sativa Mill.;
- Quantificar os aminoácidos presentes nas diferentes cultivares de castanha portuguesa;
- Obter um proteoma completo através de electroforese bidimensional do tipo IEF x SDS-PAGE de uma cultivar portuguesa de castanha;
- Identificar as sequências proteicas presentes na castanha, por intermédio de espectrometria de massa (MALDI-TOF/MS).
3. Material e Métodos
3.1. Descrição do material vegetal
Neste trabalho foram recolhidas 26 amostras de castanha, representando 21 cultivares da região Norte e Centro de Portugal presentes no banco de germoplasma do jardim botânico da Universidade Trás-os-Montes e Alto Douro.As cultivares utilizadas neste trabalho podem ser consultadas na Tabela 7, assim com as suas respectivas origens na Figura 4.
Tabela 7 – Lista de cultivares de castanhas
utilizadas neste trabalho e respectivas origens.
Cultivar Origem ‘Aveleira’ Vinhais ‘Bária’ Marvão ‘Bebim’ Campeã ‘Benfeita’ Campeã ‘Boaventura’ Vinhais ‘Côta’ Padrela
‘Demanda’ Moimenta da Beira ‘Híbrido’ Sever do Vouga
‘Judia’ 149 Vinhais
‘Judia’ 76 Carrazedo de Montenegro ‘Lada’ Carrazedo de Montenegro
‘Lamela’ Vinhais
‘Longal’ Carrazedo de Montenegro ‘Martaínha’ 86 Sernancelhe ‘Martaínha’ 23 Penedono
‘Misericórdia’ Terras de Bouro
‘Moreira’ Campeã
‘Negral’ 16 Carrazedo de Montenegro ‘Negral’ 136 Carrazedo de Montenegro
‘Passã’ Penedono
‘Rebolão’ Carrazedo de Montenegro
‘Sousã’ 8 Padrela
‘Sousã’ 149 Penedono
‘Trigueira’ Vinhais
‘Verdeal’ 81 Mêda
‘Verdeal’ 133 Vila Real Figura 4 – Distribuição geográfica das cultivares de castanhas utilizadas neste trabalho.
O facto de as amostras de castanha terem sido recolhidas do mesmo banco de germoplasma permite eliminar a contribuição diferencial das condições edafoclimáticas. As castanhas foram recolhidas e devidamente etiquetadas com os nomes das cultivares, procedendo-se posteriormente à remoção da casca, liofilização e posterior moagem. Obtendo no final uma farinha de castanha, sendo este o ponto de partida das amostras para os futuros estudos.
3.2. Análise monodimensional
3.2.1. Extracção fraccionada das proteínas de reserva da castanha
Com o objectivo de obter material biológico (proteínas) que funcionasse bem para caracterização genotípica das diferentes cultivares recorreu-se a uma técnica de extracção diferencial das proteínas de reserva de acordo com a sua solubilidade, permitindo a separação de fracções proteicas solúveis em água, solução salina, alcoólica e alcalina com posterior diálise. O protocolo de extracção utilizado por Collada e colaboradores [43] foi utilizado com algumas modificações, como por exemplo, a eliminação do processo inicial de remoção de lípidos, uma vez que a castanha apresenta baixos níveis dos mesmos (Figura 5).
Foram pesados 1,5 g de farinha de cada amostra de castanha para um falcon e foi extraído três vezes consecutivas com solução salina (NaCl 0,5M) numa relação 10:1 (v/p) a 4ºC, com agitação durante 1 hora, centrifugando-se a 30.000g, durante 20 minutos a 4ºC. No fim de cada centrifugação separou-se o sobrenadante para um novo falcon. O sobrenadante obtido na extracção salina sofreu diálise contra água para remover o sal e precipitar as globulinas, realizando novamente uma centrifugação a 30.000g, durante 20 minutos a 4ºC. Transferiu-se o sobrenadante, que continha as albuminas, para um novo falcon, permanecendo o precipitado no mesmo. Estas amostras foram desidratadas por liofilização.
O precipitado obtido na extracção salina foi extraído três vezes consecutivas com solução alcoólica (Isopropanol 55%, 2-Mercaptoetanol 2%) numa proporção de 10:1 (v/p) à temperatura ambiente, com agitação durante 1 hora, centrifugando-se a 30.000g, durante 20 minutos à temperatura ambiente. No fim de cada centrifugação o sobrenadante foi transferido para um falcon novo, sofrendo posterior diálise contra água e posterior desidratação por liofilização.
O precipitado obtido na extracção alcoólica foi extraído 3 vezes consecutivas com solução alcalina (Tetraborato de Sódio 0,012M pH 9, 2-mercaptoetanol 2%, SDS 0,5%) na proporção de 10:1 (v/p), a 60ºC, com agitação, durante 30 minutos. No fim centrifugou-se o falcon a 30.000g, à temperatura ambiente, durante 20 minutos. O sobrenadante foi transferido para um novo falcon e foi liofilizado contra água e posteriormente desidratado por liofilização. O precipitado obtido na extracção alcalina foi descartado.
Figura 5 – Extracção sequencial das proteínas de reserva da castanha (Adaptado de Collada e
colaboradores, 1986 [43]).
As fracções proteicas obtidas da liofilização foram solubilizadas em solução C com 1% DTT com uma proporção 1:50 (p/v), para serem usadas posteriormente na electroforese monodimensional.
3.2.2. Electroforese monodimensional do tipo SDS-PAGE
A electroforese monodimensional do tipo SDS-PAGE permite a análise das proteínas de reserva de alta e baixa massa molecular relativa. O protocolo usado tem por base a metodologia descrita por Singh e colaboradores [67] com algumas modificações, nomeadamente a utilização de um gel sem gradiente de concentração de acrilamida (concentração constante), o grau de porosidade e a temperatura de migração. A metodologia apresenta algumas condicionantes como a necessidade de utilizar material calibrado, introduzir o gel imediatamente após a adição dos catalisadores, ter especial atenção ao sentido da ligação dos geradores e monitorização obrigatória do momento da saída da frente de migração do gel.
A montagem de placas para electroforese foi realizada mediante a metodologia descrita por Igrejas [68]. Os elementos envolvidos na montagem e necessários para a migração encontram-se ilustrados na Figura 6.
Figura 6 – Placas e elementos do sistema da tina de electroforese vertical. A) Tampa da tina; B)
Tina de electroforese; C) Bandeja superior; D) Nível; E) Pente; F) Parafusos; G) Espaçador; H) Braçadeira; I) Vidros; J) Suporte; K) Dispositivo de refrigeração. (Fonte: Igrejas, 1996 [69]).
Inicialmente procedeu-se à preparação de um gel SDS-PAGE de 16x18 cm, 1 mm de espessura e de 15 poços. A Tabela 8 apresenta as quantidades necessárias das soluções para a preparação do gel de separação para as proteínas de reserva de alta e baixa massa molecular relativa, tendo sido utilizado a concentração de T: 12,52% e C: 0,97%.
Após a mistura dos primeiros quatro reagentes, a solução foi depositada num kitasato selado e com a ajuda de uma bomba de vácuo procedeu-se à desgaseificação durante 10 minutos. Após a desgaseificação adicionaram-se os restantes reagentes pela ordem apresentada na Tabela 8. O TEMED deve ser adicionado à solução com a ponta da micropipeta e ressuspendido, para uma melhor polimerização. A solução foi transferida para um copo e com o auxílio de uma seringa depositou-se nas placas de electroforese, deixando uma margem de 3 cm até ao topo, para que posteriormente seja também depositado o gel de concentração. Após a deposição da mesma, são adicionados 180 μl de butanol ao longo da sua superfície, de modo a prevenir a formação de novas bolhas de ar. Enquanto ocorria a polimerização do gel de separação, ordenaram-se as amostras por ordem de inserção e preparou-se um registo para as amostras e condições de migração. Verificou-se a polimerização do gel e preparou-se o gel de concentração a ser adicionado superiormente ao gel de separação (T = 2,88 e C = 1,42%) (Tabela 9), em que os passos necessários para a realização deste gel foram idênticos ao gel de separação.
Antes que ocorra a polimerização do mesmo, foi introduzido um pente de 15 poços. Após a polimerização do gel, retirou-se o pente e lavaram-se os poços três vezes. Antes de proceder à deposição das amostras, estas foram pré aquecidas na estufa a 70ºC e centrifugadas durante 5 minutos, a 10.000 rpm. Depositou-se aproximadamente 10 μl de amostra por poço. Colocou-se a bandeja superior na tina de electroforese. Cobriu-se a bandeja com tampão de electroforese, ligando-a depois a uma fonte de alimentação. As condições de migração são de 30 mA/gel a 14 °C. A migração foi parada no momento em que a frente de migração saiu das placas.
Tabela 8 – Preparação dos géis de separação (T: 12,52% e C: 0,97%).
1 gel 2 géis 4 géis 6 géis
Acrilamida 40% 7,655 ml 15,31ml 30,62 ml 45,93 ml
Bisacrilamida 2% 1,5 ml 3 ml 6 ml 9 ml
Água bidestilada 5,25 ml 10,5 ml 21 ml 31,5 ml
Tampão Tris-HCL pH 8.8 9,4 ml 18,8 ml 37,6 ml 56,4 ml Desgaseificação (com bomba de vácuo)
SDS 10% 0,25 ml 0,5 ml 1 ml 1,5 ml
APS 1% 0,625 ml 1,25 ml 2,5 ml 3,75 ml
TEMED 12,5 ul 25 ul 50 ul 75 ul
![Figura 2 – Os dois eventos migratórios do género Castanea com base na análise filogenética para a região trnL-trnF de DNA cloroplastidial (Adaptado de Abreu, 2007 [7])](https://thumb-eu.123doks.com/thumbv2/123dok_br/15772709.1076102/23.892.192.741.119.526/figura-eventos-migratórios-castanea-análise-filogenética-cloroplastidial-adaptado.webp)
![Figura 3 – Distribuição geográfica da produção de castanha no ano 2014 (Adaptado de FAOSTAT, 2017 [12])](https://thumb-eu.123doks.com/thumbv2/123dok_br/15772709.1076102/24.892.132.765.695.1048/figura-distribuição-geográfica-produção-castanha-ano-adaptado-faostat.webp)
![Tabela 2 – Produção de castanha na Europa no ano 2014 (Adaptado de FAOSTAT, 2017 [12])](https://thumb-eu.123doks.com/thumbv2/123dok_br/15772709.1076102/25.892.128.764.451.682/tabela-produção-de-castanha-europa-ano-adaptado-faostat.webp)

![Tabela 4 - Composição centesimal de macronutrientes, fibras e valor energético da castanha portuguesa (g/100g) (Adaptado de Souza e colaboradores, 2014 [19])](https://thumb-eu.123doks.com/thumbv2/123dok_br/15772709.1076102/27.892.137.747.752.1067/composição-centesimal-macronutrientes-energético-castanha-portuguesa-adaptado-colaboradores.webp)
![Tabela 5 - Distribuição geográfica das diferentes cultivares Portuguesas de castanha (Adaptado de Ferreira-Cardoso e Pimentel-Pereira, 2007 [8])](https://thumb-eu.123doks.com/thumbv2/123dok_br/15772709.1076102/31.892.124.771.146.1051/distribuição-geográfica-diferentes-cultivares-portuguesas-adaptado-ferreira-pimentel.webp)
![Figura 5 – Extracção sequencial das proteínas de reserva da castanha (Adaptado de Collada e colaboradores, 1986 [43])](https://thumb-eu.123doks.com/thumbv2/123dok_br/15772709.1076102/44.892.149.751.348.759/figura-extracção-sequencial-proteínas-castanha-adaptado-collada-colaboradores.webp)

