Universidade de Trás-os-Montes e Alto Douro
Sistema Olfativo do Cão
Estabelecimento de uma Metodologia de Abordagem
à Lâmina Crivosa do Cão
Tese de Dissertação de Mestrado Integrado em Medicina
Veterinária
Rui Damásio Alvites
Orientador:
Professor Doutor Artur Severo Proença Varejão
Universidade de Trás-os-Montes e Alto Douro
Coorientador:
Professor Doutor Bruno Jorge Antunes Colaço
Universidade de Trás-os-Montes e Alto Douro
ii
Universidade de Trás-os-Montes e Alto Douro
Sistema Olfativo do Cão
Estabelecimento de uma Metodologia de Abordagem
à Lâmina Crivosa do Cão
Tese de Dissertação de Mestrado Integrado em Medicina
Veterinária
Rui Damásio Alvites
Orientador:
Professor Doutor Artur Severo Proença Varejão
Universidade de Trás-os-Montes e Alto Douro
Coorientador:
Professor Doutor Bruno Jorge Antunes Colaço
Universidade de Trás-os-Montes e Alto Douro
Composição do Júri:
Presidente: Professora Doutora Cristina Maria Teixeira Saraiva
Orientador: Professor Doutor Artur Severo Proença Varejão
Arguente: Professor Doutor Carlos Alberto Antunes Viegas
iii
DECLARAÇÃO
NOME: Rui Damásio Alvites
C.C: 13723523
TELEMÓVEL: (+351) 938431195
CORREIO ELECTRÓNICO: ruialvites@hotmail.com
DESIGNAÇÃO DO MESTRADO: MESTRADO INTEGRADO EM
MEDICINA VETERINÁRIA
TÍTULO DA DISSERTAÇÃO DE MESTRADO EM MEDICINA
VETERINÁRIA: Estabelecimento de uma Metodologia de Abordagem à
Lamina Crivosa do Cão
ORIENTADORES:
Professor Doutor Artur Severo Proença Varejão
Professor Doutor Bruno Jorge Antunes Colaço
ANO DE CONCLUSÃO: 2015
DECLARO QUE ESTA DISSERTAÇÃO DE MESTRADO É
RESULTADO DA MINHA PESQUISA E TRABALHO PESSOAL E DAS
ORIENTAÇÕES DOS MEUS SUPERVISORES. O SEU CONTEÚDO É
ORIGINAL E TODAS AS FONTES CONSULTADAS ESTÃO
DEVIDAMENTE MENCIONADAS NO TEXTO, E NA BIBLIOGRAFIA
FINAL. DECLARO AINDA QUE ESTE TRABALHO NÃO FOI
APRESENTADO EM NENHUMA OUTRA INSTITUIÇÃO PARA
OBTENÇÃO DE QUALQUER GRAU ACADÉMICO.
VILA REAL, 28 DE Novembro de 2015.
Rui Damásio Alvites
iv
À minha mãe Ao meu pai À Minha irmã A todos os que entraram na minha vida e dela não saíra
v
Agradecimentos
A conclusão da dissertação de mestrado é o culminar de um percurso académico longo, marcado pela sua dureza e pela complexidade, mas também recheado de momentos memoráveis amizades profundas e pessoas inesquecíveis. Por sua vez, todo este percurso surge na sequência de uma vida plena de figuras importantes que me permitiram chegar a este momento, sendo a pessoa que sou. A todas elas, e por mais que seja lugar-comum, é preciso agradecer o contributo que tiveram na minha vida, no meu crescimento, na minha identidade.
Começo por agradecer à minha mãe, a pessoa mais importante da minha vida, por todo o amor e companheirismo com que sempre me agraciou, por ser uma amiga acima de todas as relações de parentesco, por me ter ensinado todos os valores por que hoje me rejo, por ser um exemplo de força e perseverança e por servir de âncora inquebrável em todos os momentos da minha vida. Também ao meu pai por me ter ensinado o valor do trabalho, a importância da dedicação à família e por me ter mostrado ao longo da vida que a manifestação dos afetos pode ser feita de várias formas, todas elas com igual valor.
Á minha irmã, companheira de mil e uma aventuras e cumplicidades, por ter iluminado a minha infância e adolescência e por ter mantido vivo o meu lado de criança por todos estes anos. Ao meu tio Augusto, pelo amor incondicional que sempre demonstrou, por me ter ensinado tanto e tão bem, por me ter mostrado o valor do trabalho e da palavra na construção de uma personalidade forte e inquestionável, pelos tantos sonhos realizados ao seu lado. Á minha avó por ter preenchido a minha memória com tantas boas lembranças. Às minhas primas, Elsa e Alison, por terem crescido comigo, pelos momentos partilhados, pelas piadas que só nos percebemos, por serem acima de tudo duas irmãs que vivem longe. A toda a família Damásio por serem quem são, por me terem criado tão bem, simplesmente por existirem.
A todos os amigos de infância que cresceram e viveram aventuras ao meu lado, em particular à Carminda, a eterna vizinha, tão presente nas minhas primeiras memórias que parece não ter existido um tempo antes de a conhecer. Ao João, por me ter acompanhado durante o crescimento, pelos vícios que partilhamos e por continuar a ser o amigo que sempre foi, passados tantos anos. Ao Hugo e ao Rui, amigos de sempre e para sempre, por todas as aventuras vividas, as memórias partilhadas, as piadas privadas que nos fazem rir sem parar, por todos os reencontros, ano após anos, em que parece não ter passado um só dia desde a separação.
Chegado à universidade, um agradecimento conjunto a todos aqueles que percorreram a estrada da vida académica ao meu lado e que fizeram deste período a fase inesquecível a que chamamos os melhores anos da nossa vida. Especificando, agradeço à Rute, a primeira a quem abri o meu coração no meio de tanta gente estranha, por me ter ouvido, aconselhado e apoiado, por ter ido e voltado, pelo humor corrosivo que partilhámos e pelas conversas tão boas, que embora cada vez mais raras são sempre momentos de prazer. Á
vi
Joana, companheira desta longa jornada, por me perceber tão bem, por me ouvir sempre que necessário, por ter vivido ao meu lado aventuras dentro e fora da faculdade, pela dura história de vida e por ser, agora mais que nunca, um exemplo de profissionalismo e dedicação. À Rita, a amiga cujo humor é tão igual ao meu que jamais poderei partilhar com mais ninguém as piadas e a imagens que tanto nos fizeram rir, pelos desabafos, confissões e intimidade, pelos sonhos partilhados e conspirações arquitetadas que mais ninguém podia ouvir, por tudo aquilo que passamos juntos ao longo destes anos. Á Flávia, amiga desde sempre e possuidora de uma mente tão brilhante que continua a ser assustador pensar nas suas potencialidades, por ter percorrido toda esta jornada comigo e ter continuado ao meu lado depois disso, pelas mil e uma ajudas, concelhos e desenrasques, pelas inúmeras sessões de cinema e jantares aqui e acolá, por tudo o que me ensinou e o que ainda ensinará. Ao Neca, uma fonte inesgotável de conhecimento geral, pelos gostos culturais partilhados, pela calma e placidez em todos os momentos e por ter sempre algo de novo a ensinar em cada conversa.
A todos os professores que me ensinaram e me deram ferramentas para construir o meu conhecimento, em especial ao professor Artur Varejão pelo exemplo de profissionalismo e polivalência e por me ter feito apaixonar pelo mundo da neurologia e ao professor Bruno Colaço pelo seu companheirismo, por ter ouvido sem relutâncias as minhas ideias e me ter apoiado em cada fase deste projeto, sempre com novas ideias e com uma palavra de encorajamento a cada nova reunião.
Por fim, e chegando à fase de elaboração da própria dissertação, um especial agradecimento ao HVP e a tudo o que ele representou ao longo deste ano, aos funcionários, estagiários e médicos que fizeram desta experiência a mais intensa fase da minha vida, à família que aqui construí, aos conhecimentos que adquiri, às mil e uma histórias que levo para contar. Á Sara, não só por todos os momentos partilhados e pela preocupação desmensurada, mas também pelo trabalho fotográfico exímio sem o qual esta dissertação nunca teria sido concluída.
E claro, um sentido e especial agradecimento ao grupo de estagiários que viveram comigo esta fase e se tornaram na minha segunda família. Por todos os momentos, os jantares, as saídas, as viagens, as conspirações, a zangas, os risos, os momentos… por terem entrado na minha vida, a terem virado do avesso e terem feito de mim uma pessoa melhor. Especificando, mas sem fazer jus a tudo o que haveria para dizer, á Filipa e as sessões de conspiração, mal dizer e a todos os inimigos que partilhámos; á Catarina por ter trazido o “Brasiu” até Portugal e me ter ensinado a procurar o ganso e nunca me perder na multidão, à Daniela por me ter mostrado todo um novo mundo nipónico e me ter deixado ser o seu filhote mesmo depois das feias discussões; à Paula por ser uma fada, me ter apresentado a querida Bambi e ter trazido tanta fantasia aos momentos mais tensos dos meus dias, à Carolina por todos os momentos de cumplicidade, pelo exemplo de profissionalismo e perseverança e por me ter ensinado que “Nâ sê” e “Moce” também são palavras, de novo à Flávia por me ter acompanhado em mais esta fase da minha vida e se ter tornado num pequeno monstrinho que ainda assim continuo a adorar, ao Filipe pelo companheirismo, pelas mil conversas e confissões, por estar aqui sempre que necessário, por se ter tornado num irmão sem qual nada disto teria tido o mesmo significado.
vii
Resumo
A lâmina crivosa do osso etmóide contém um grande número de forâmenes que permitem a passagem dos feixes nervosos do par craniano I desde o epitélio olfativo até ao bulbo olfativo, garantindo assim a transmissão da informação olfativa recolhida a nível da cavidade nasal e a sua integração a nível cerebral. Apesar da sua estreita relação e importância na capacidade olfativa, não há registo de uma abordagem completa a esta estrutura no cão no que toca às suas características morfológicas e à sua influência no olfato canino.
O objetivo deste trabalho é a criação de uma metologia de análise qualitativa e quantitativa das características da lâmina crivosa no cão, permitindo assim mensurar a sua área, contabilizar o seu número de forâmenes e ainda determinar a área ocupada pelos mesmos na sua superfície, além de estabelecer relações com outros componentes e medidas do crânio do animal através de métodos combinados de observação direta, captação e tratamento de imagem. Para tal foram utilizados seis crânios submetidos à mensuração de nove medidas craniométricas previamente estabelecidas para permitir um estudo estatístico posterior da sua relação com as dimensões e número de forâmenes da lâmina crivosa.
A aplicação da metodologia criada permitiu concluir que a lâmina crivosa não é morfologicamente idêntica em todos os crânios, não se observando simetria entre as suas duas metades nem quanto à forma, nem em dimensões, nem tampouco no número e distribuição de forâmenes na sua superfície. A relação entre as três variáveis em estudo na lâmina crivosa não é forte de igual forma: embora as três apresentem correlações positivas entre si, a única relação verdadeiramente previsível estabelece-se entre a área total da lâmina crivosa e a área ocupada pelos forâmenes na mesma. As correlações entre o número de forâmenes e as outras duas variáveis são mais baixas.
As relações estabelecidas entre as variáveis da lâmina crivosa e as medidas craniométricas selecionadas são, na sua maioria, explicáveis à luz dos conhecimentos atuais sobre morfometria do crânio do cão e sua influência na fisiologia olfativa canina, abrindo portas para um grande número de possibilidades a explorar no futuro.
viii
Abstract
The cribriform plate of the ethmoid bone contains a large number of foramina that allow the passage of the bundles of the cranial nerve I from the olfactory epithelium to the olfactory bulb, thus assuring transmission of olfactory information collected at the level of nasal cavity and its integration in the brain. Despite its close relation and importance regarding olfactory capacity, there is no record of complete approaches to this structure in the dog relative to their morphological characteristics and its influence in the canine olfaction
The purpose of this work is the creation of a methodology that qualifies and quantifies the canine cribriform plate features that allow measuring its area, count the number of foramina and also determine the area occupied by them on its surface, as well as establish relationships with other components and measures of the animal skull through combined methods of direct observation, image capture and processing.For this purpose, six skulls were used and submitted to a mensuration of nine craniometric measures, previously established, to allow the further statistical study of its relations with the cribriform plate dimensions and its foramina number.
The application of this established methodology allowed to conclude that the cribriform plate is not morphologically identical in every skull, having no symmetry between the two halves, neither in shape or dimensions, not even in the number and distribution of foramina on its surface. The relation between the three variables studied in the cribriform plate is not equally strong: although the three show positive correlation with each other, the only predictable relation is established between the area of the cribriform plate and the area occupied by the foramina in its surface. The correlation between the number of foramina and the other two variables is weaker.
The relations established between the variables of the plate and the craniometric measures selected are, mostly, explained in the light of current knowledge about dog’s skull morphology and its influence in the canine olfactory physiology, opening doors to a large number of possibilities in the future
ix
Índice Geral
Dedicatória ... iv Agradecimentos ... v Resumo ... vii Abstract ... viii Lista de Figuras ... xiLista de Tabelas ... xii
Lista de Abreviaturas e Siglas ... xiii
Capítulo 1. Revisão Bibliográfica ... xiv
1 Introdução ... 1
1.1 Sentido do Olfato ... 2
1.2 Sistema Olfativo ... 4
1.2.1 Morfologia do Sistema Olfativo ... 4
1.2.1.1 Sistema Olfativo Principal ... 4
1.2.1.1.1 Cavidade Nasal ... 5
1.2.1.1.2 Nervo Olfativo (Par Craniano I) ... 12
1.2.1.1.3 Rinencéfalo ... 14
1.2.1.2 Sistema Olfativo Acessório ... 19
2 Fisiologia do Sistema Olfativo ... 21
2.1 Fisiologia do Sistema Olfativo Principal ... 21
2.1.1 Fluxo de Ar Nasal ... 23
2.1.2 O Ato de Farejar ... 25
2.1.3 Dinâmica Extranasal de Fluidos ... 25
2.1.4 Dinâmica Intranasal de Fluidos ... 26
2.1.5 Deposição e Captação das Moléculas Odoríferas no Recesso Olfativo………..27
2.1.6 Difusão das Moléculas Odoríferas através da Mucosa Olfativa ... 28
2.1.7 Ligação das moléculas Odoríferas aos Recetores Olfativos ... 29
2.1.8 Eliminação dos Complexos PLO-Odorante ... 29
2.1.9 Vias de Transdução ... 30
2.1.10 Vias Olfativas ... 32
x
3 Alterações do Sistema Olfativo ... 38
3.1 Exame Neurológico aplicado ao Par Craniano I ... 39
3.2 Diminuição da renovação da Mucosa Nasal ... 40
3.3 Alterações da Lâmina Crivosa ... 43
3.4 Doenças Neurodegenerativas ... 44
Capítulo 2. Estabelecimento de uma Metodologia de Abordagem à Lâmina Crivosa do cão……….……xv
1 Objetivos ... 50
2 Materiais e Métodos ... 51
2.1 Crânios ... 51
2.2 Mensuração Craniana Inicial ... 51
2.3 Análise da Lâmina Crivosa ... 54
2.4 Análise Estatística ... 59
3 Resultados ... 59
3.1 Medidas Craniométricas ... 59
3.2 Caracterização Morfológica da Lâmina Crivosa ... 61
3.3 Análise da Lâmina Crivosa ... 62
3.4 Correlação entre as Características da Lâmina Crivosa e as Medidas Craniométricas ... 65
4 Discussão ... 68
4.1 Estabelecimento da Metodologia de Abordagem à Lâmina Crivosa do Cão ... 68
4.2 Observações Qualitativas e Quantitativas da Lâmina Crivosa ... 69
4.3 Relação entre as três Variáveis da Lâmina Crivosa ... 71
4.4 Assimetria entre as Duas Metades da Lâmina Crivosa ... 72
4.5 Relação entre as três Variáveis da Lâmina Crivosa e as Medidas Craniométricas. 73 5 Trabalho Futuro ... 77
xi
Lista de Figuras
Figura 1. Representação esquemática dos componentes que constituem o SO principal
canino………..…....5
Figura 2. Corte sagital da CN do cão, evidenciando as conchas e meatos nasais………...7
Figura 3. Secção transversal do crânio num plano caudal à LC………8
Figura 4. Vista lateral do septo nasal num cão adulto……...………...…………...10
Figura 5. Esquema representativo do epitélio olfativo………...………12
Figura 6. Corte sagital da CN evidenciando a distribuição dos nervos na mucosa septal e o OV………...………..14
Figura 7. Diagrama esquemático representando as camadas histológicas do BO, as interações entre as diferentes células bulbares e os circuitos neurais básicos do BO……….………16
Figura 8. Vista ventral do cérebro ………..………...18
Figura 9. Recesso olfativo no cão………..………23
Figura 10. Vias aéreas do cão ………...………...…..24
Figura 11. Dinâmica extranasal de fluidos no cão………26
Figura 12. Ilustração esquemática do fenómeno de transporte de moléculas odoríferas desde o ar inspirado até aos recetores olfativos localizadas na superfície epitelial……….……….28
Figura 13. Esquema representativo da teoria da ligação segundo a forma e ativação de sinal respetivo ……….………..29
Figura 14. Esquema representativo da transdução sensorial dos sinais olfativos…………...…31
Figura 15. Representação esquemática das vias olfativa……….……….34
Figura 16. Esquema da transdução nas células recetoras vomeronasais, nos recetores V1R……….…..38
Figura 17. Aspeto normal dos ectoturbinados na CN; Assimetria entre os turbinados das duas CN………...………..41
Figura 18. LC humanas……….……….43
Figura 19. Vista ventral e vista dorsal de crânio de cão com representação das medidas craniométricas mensuradas………...52 Figura 20. Identificação da largura (L) e comprimento (C) usados na determinação do IS; Corte de um dos crânios com serra da fita vertical; Identificação do local de corte do crânio;
xii
observação do aspeto cranial da cavidade craniana e superfície caudal da LC após corte do crânio……….54 Figura 21. Imagem original da LC; Aplicação da ferramenta “laço”; Imagem final da lâmina isolada………..……….55 Figura 22. Criação de perfil de calibração………..55 Figura 23. Imagem da LC após segmentação……….………56 Figura 24. Contagem automática do número de forâmenes; Pormenor da tabela de resultados ………..……...57 Figura 25. Exemplo de tabela de análise estatística ………..….57 Figura 26. Divisão do número de forâmenes em duas categorias; Tabela de resultados ………...………58 Figura 27. Imagem de perfil da LC após ser submetida à ferramenta “exposição”……….58 Figura 28. Estudo idêntico ao realizado para a LC como um todo, aplicado apenas à metade direita………..………….….59 Figura 29. Foto de LC normal e foto de LC segmentada………61 Figura 30. Assimetria entre as metades esquerda e direita da LC……….……….62 Figura 31. Gráficos de regressão linear da relação das três variáveis em estudo………..63 Figura 32. Fotografias das LC dos crânios 1 e 3……….…..……..70
Lista de Tabelas
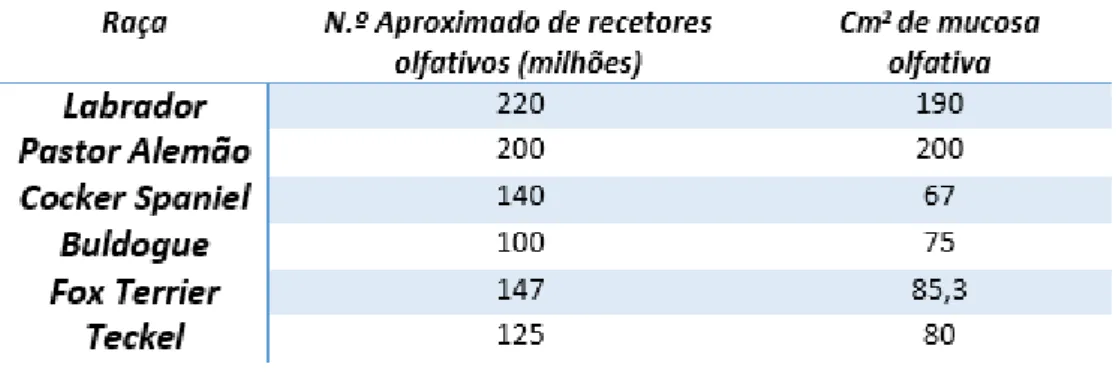
Tabela 1. Relação entre a raça/dimensões do animal, número aproximado de recetores olfativos na CN e área ocupada pela mucosa olfativa na CN………...……….22 Tabela 2. Medidas craniométricas obtidas nos seis crânios em estudo …….……….60 Tabela 3. Resultados obtidos a partir da análise das LC dos seis crânios em estudo………….62 Tabela 4. Resultados obtidos a partir da análise de cada uma das metades da LC…………..…64 Tabela 5. Relação estatística entre cada uma das medidas craniométricas e os valores da LC e seus forâmenes………...…….65 Tabela 6. Relação estatística entre cada uma das medidas craniométricas e o ratio entre a área total ocupada pelos forâmenes na lâmina e a área total da LC………67
xiii
Lista de Abreviaturas e Siglas
BO – Bulbo OlfativoBOA – Bulbo Olfativo Acessório cAMP – AMP- cíclico
CN – Cavidade Nasal
CV – Coeficiente de Variação DAG – Diacilglicerol
DT – Distância Total
ECAV – Escola de Ciências Agrárias e Veterinárias da UTAD RAPG – Recetores acoplados à Proteínas G
IP3 – Trifosfato de Inositol IS – Índex Sexual
LC – Lâmina Crivosa MO – Molécula Odorífera NO – Nervo Olfativo OV – Órgão Vomeronasal
PLO – Proteínas de Ligação a Odorantes SNC – Sistema Nervoso Central
SO – Sistema Olfativo SV – Sistema Vomeronasal
TRLC2 - Canais iónios Recetores de Potencial Transitório – Média
σ – Desvio-padrão
xiv
Capítulo 1
1
1. Introdução
Os mamíferos têm a capacidade de reconhecer um elevado número de sinais químicos no seu ambiente envolvente, o que influencia grandemente o seu comportamento e lhes confere informação ambiental essencial à sua sobrevivência. Nos organismos superiores, embora a informação química ambiental seja recolhida tanto pelo olfato como pelo paladar, muitas espécies dependem do olfato como principal sistema de receção química. Assim, o Sistema
Olfativo (SO) nos mamíferos regula uma grande variedade de funções integrativas como
homeostasia, respostas emocionais (como ansiedade, medo e prazer), reprodução (comportamento sexual e maternal) e interações sociais (reconhecimento de animais da mesma espécie, família, clã ou invasores) (LEDDO et al., 2005) e a sua utilização na deteção de doenças, contaminação alimentar, agentes nocivos e até em potenciais aplicações comerciais tem chamado a atenção e rendido um aumento do investimento na investigação em torno do olfato e das suas potencialidades (SANKARAN et al., 2012).
O cão tem sido largamente utilizado desde os primórdios em tarefas que exigem o uso das suas capacidades olfativas como seja a caça, deteção de drogas e explosivos e identificação de doenças específicas (principalmente neoplasias e doenças endócrinas) (GRANDJEAN & HAYMANN). Apesar de tudo, ainda não conseguimos quantificar precisamente a acuidade olfativa desta espécie nem a percentagem de capacidade olfativa que estes animais usam em cada uma destas atividades. Surgem dúvidas quanto aos fatores anatómicos e fisiológicos que distinguem diferentes raças de cães e a sua apetência para determinadas tarefas e também quanto à experiencia olfativa de um cão não treinado que, apesar de ter um SO tão preparado como um cão submetido a treino, não demonstra as mesmas capacidades que estes perante as mesmas situações e estímulos (HOROWITZ et al., 2013).
A Lâmina Crivosa (LC) (lamina cribrosa) do osso etmóide contém diversos forâmenes que permitem a passagem dos filamentos do par craniano I desde o epitélio olfativo até ao
Bulbo Olfativo (BO) (Bulbus olfactorius), garantindo a transmissão dos estímulos olfativos
desde a Cavidade Nasal (Cavum nasi) (CN), até ao cérebro (KALMEY et al., 1998). O impacto desta estrutura na capacidade olfativa do cão e sua influência na variabilidade detetada entre diferentes animais e raças ainda não foi determinada. Estudar as caraterísticas morfológicas da LC, as relações estabelecidas entre as variáveis desta estrutura, entre estas e outras medidas
2
craniométricas e a criação de um protocolo de abordagem à LC canina são os objetivos do presente trabalho.
1.1 Sentido do Olfato
A perceção sensorial é o processo através do qual a informação do ambiente exterior é reformatada para um estado assimilável no meio interno. A origem da capacidade de detetar moléculas químicas exteriores surgiu e tem vindo a evoluir desde os procariotas, tendo progredido para quatro modalidade distintas nos vertebrados: o SO principal, o SO acessório, o sistema gustativo e o chamado sistema químico comum associado maioritariamente aos neurónios sensoriais do nervo trigémio (LEDDO et al., 2005). Basicamente, quatro componentes são obrigatórios em todos os SO, sendo eles: a presença de proteínas que se liguem às moléculas odoríferas; a presença de recetores específicos associados a proteínas G que funcionem como recetores de moléculas odoríferas; o desencadear de uma cascata de transdução de sinal de duas fases e a presença de estruturas funcionais a nível central que recebam as informações das vias olfativas (HILDEBRAND & SHEPHERD, 1997).
O SO é o sistema sensorial químico responsável pela capacidade de captar, processar e identificar moléculas químicas com origem numa fonte distante (em contraste com o sistema gustativo onde se exige um contacto direto com a fonte da molécula química) (LEDDO et al., 2005), enquanto Olfato é a função do SO através da qual os odores são identificados (SOUDRY
et al., 2011). O conjunto de órgãos envolvidos na olfação constitui o SO e embora possa variar
estruturalmente em diferentes animais, de forma geral os neurónios sensoriais envolvidos estão sempre localizados perto da superfície corporal (RASTOGI, 2007). O SO é ainda hoje o menos compreendido e explorado de entre os chamados cinco sentidos sensoriais. Denomina-se de
odorante ou Molécula Odorífera (MO) todo o composto volátil capaz de desencadear uma
resposta olfativa, enquanto odor é a sensação resultante do estímulo dos órgãos do SO. Os odorantes e a sua perceção desempenham funções importantes na fisiologia e comportamento animal, tendo influência sobre fatores como a estimulação do apetite, deteção de perigos eminentes e interações sociais. Os odorantes detetados podem ter um impacto significativo na etologia do animal, nomeadamente criando associações com experiências passadas que levam ao estabelecimento de uma escala de categorização dos odores, classificados como agradáveis, desagradáveis ou indiferentes consoante as situações previamente vividas. O impacto que os odores podem ter sobre o animal varia de indivíduo para indivíduo (BARRET et al., 2010).
3
Animais com olfato desenvolvido (cão e a maior parte dos animais domésticos) são denominados de macrosmáticos; animais com olfato deficitário (homem, outros primatas e alguns mamíferos aquáticos) são denominados microsmáticos; por fim, animais sem capacidade olfativa (a maior parte dos mamíferos aquáticos) são anosmáticos (REECE, 2009). Todos os vertebrados possuem um SO principal, sendo que a maior parte dos vertebrados terrestres apresenta ainda o Sistema Vomeronasal (SV) (HILL et al., 2012).
No processo evolutivo das formas de vida primitivas, os corpos celulares das células nervosas migraram em sentido central no organismo, sendo que apenas os prolongamentos nervosos se mantiveram em posição periférica. Esta posição central dos corpos celulares resulta numa proteção extra contra lesões e destruição por fatores externos e a manutenção do corpo celular garante que, em caso de agressão contra os prolongamentos nervosos, possa ocorrer regeneração em diferente extensão. Esta migração não se verificou nos corpos celulares do par craniano I ou Nervo Olfativo (NO) (Nn. olftorii), encontrando-se estes a nível da membrana mucosa da CN, na chamada região olfativa. Esta localização periférica torna estes corpos celulares nervosos mais suscetíveis a destruição por agressões externas, principalmente de origem inflamatória. Assim, sob determinadas condições de agressão, animais macrosmáticos e microsmáticos podem tornar-se temporária ou definitivamente microsmáticos e anosmáticos (REECE, 2009).
Enquanto espécie, o Cão (Canis familiaris) tem sido alvo de uma crescente atenção por parte da investigação biomédica na medida em que partilha com o homem uma variedade genética e clínica assinalável. A CN do cão tem sido alvo de diversos estudos que permitem hoje conhecer bem a sua anatomia e fisiologia e até os padrões dos fluxos de ar através da mesma durante os movimentos respiratórios (BARRIOS et al., 2014). Sabe-se que possui uma CN particularmente desenvolvida e considerada como um dos melhores detetores de odorantes da natureza (LAWSON et al., 2012) e, portanto, uma elevada acuidade olfativa (CRAVEN et
al., 2009). A investigação no âmbito da identificação, diagnóstico e prognóstico de tumores
nasais, os estudos sobre a mucosa da CN e a sua peculiar manutenção da população de células basais e ensaios clínicos sobre implantação de células derivadas de culturas autólogas de mucosa olfativa e o seu impacto na atenuação dos efeitos em traumatismos medulares agudos são áreas sob especial atenção por parte da comunidade científica nos últimos anos (BARRIOS
4
1.2 Sistema Olfativo
Nos ambientes terrestes, os sinais químicos podem ter origem volátil ou não volátil, obrigando os vertebrados a possuir dois SO anatomicamente e funcionalmente distintos: o SO principal para detetar compostos voláteis e o SO acessório ou SV para detetar compostos não voláteis (embora não exclusivamente) (QUIGNON et al., 2005). O SO principal ou comum funciona como sensor ambiental, sendo utilizado pelo animal na procura de alimento, deteção de predadores e presas e marcação territorial e notabilizando-se pela sua amplitude e poder de discriminação. Este sistema desenvolveu-se sob o pressuposto de que é impossível prever, a
priori, com que tipo de moléculas irá contactar, sendo necessário manter uma matriz sensorial
indeterminada, mas ainda assim precisa. O SO acessório ou vomeronasal desenvolveu-se com a função reprodutiva específica de identificação de um parceiro recetivo, uma função complexa o suficiente para garantir a presença de um sistema independente. Este sistema especializou-se no reconhecimento de sinais olfativos espécie-específicos produzidos por um género e reconhecidos pelo outro e interpreta informações não só sobre a localização mas também sobre o estado reprodutivo e disponibilidade do possível parceiro sexual. Além disso também desemprenha funções importantes no comportamento social, marcação territorial, agressividade e amamentação (FIRESTEIN, 2001).
1.2.1 Morfologia do Sistema Olfativo
1.2.1.1 Sistema Olfativo Principal
Os componentes do SO principal estão representados de forma resumida na Figura 1. Estes podem basicamente ser divididos em duas grandes regiões: a CN e o rinencéfalo (Rhinencephalon), na cavidade craniana. Na primeira encontramos os recetores olfativos, a nível da mucosa nasal, onde se dá o primeiro contacto com as MO; a nível do rinencéfalo encontramos o BO que recebe a informação olfativa proveniente da mucosa olfativa e tem a função de a transmitir às diferentes regiões do córtex olfativo. Estas regiões de integração da informação olfativa estabelecem ainda diversas vias de comunicação entre si e com outras regiões do encéfalo (JIAL et al., 2014). Cada uma destas regiões e componentes são explorados e descritos de seguida, de forma sucinta.
5
1.2.1.1.1 Cavidade Nasal
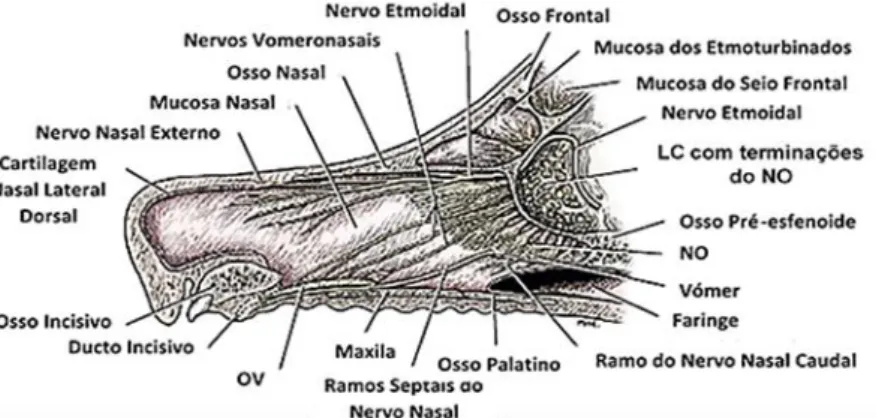
A CN do cão (Figura 2) é a porção mais rostral das vias respiratórias. Estende-se desde as narinas (Nares) até às coanas (Choanae), sendo dividida pelo septo nasal (Septum nasi) em duas vias aéreas simétricas bilateralmente. Este septo nasal é constituído por uma porção óssea (Pars ossea), uma porção cartilagínea (Cartilago septi nasi), e ainda uma porção membranosa (Pars membranacea). Ambas as cavidades apresentam três regiões anatómicas bem definidas:
vestíbulo nasal (Vestibulum nasi), a porção mais rostral, revestido por pele e contendo folículos
pilosos e glândulas sebáceas (SANKARAN et al., 2012); região respiratória e região olfativa. O vestíbulo é responsável por aquecer ou arrefecer, humidificar e filtrar o ar inspirado antes de este penetrar no trato respiratório inferior. Tendo em conta a esparsa vascularização encontrada no vestíbulo nasal, pouco ar é condicionado nesta região, podendo ainda assim ocorrer alguma filtração. Assim, o vestíbulo nasal é responsável principalmente por distribuir o ar inspirado para dentro da CN e também para direcionar o ar expirado (CRAVEN et al., 2007). O osso
etmoide (Os ethmoidale) localiza-se no limite caudal da CN e está separado da cavidade craniana pela LC, de posição transversal. O labirinto etmoidal (Labyrinthus ethmoidalis) origina-se a partir da parede dorsal e lateral da lâmina externa do osso frontal e é constituído por finas lamelas ósseas, os etmoturbinados (Ethmoturbinalia), entre os quais surgem espaços
Figura 1: Representação esquemática dos componentes que constituem o SO principal canino (adaptado de JIAL
6
denominados de meatos etmoidais (Meatus ethmoidales). Os etmoturbinados, maiores e de posição interna, são chamados de endoturbinados (Endoturbinalia), enquanto os mais pequenos e externos recebem o nome de ectoturbinados (Ectoturbinalia), surgindo estes numa relação de 4/6 no cão (KÖNIG & LIEBICH, 2004).
Por sua vez, as conchas nasais (Conchae nasales) são estruturas convolutas cartilagíneas ou ossificadas, cobertas por mucosa nasal, que ocupam a maior parte das CN. Estas constituem a concha nasal dorsal (Concha nasalis dorsalis) ou nasoturbinado, concha
nasal média(Concha nasalis media) e concha nasal ventral (Concha nasalis ventralis) ou
maxiloturbinado (EVANS & LAHUNTA, 2013). Do ponto de vista funcional, é
essencialmente a complexa estrutura do maxiloturbinado que garante uma elevada área de superfície para transferências de calor e humidade. No cão esta concha ventral é do tipo ramificado garantindo uma ainda maior área de superfície, sendo que a tortuosidade entre os seus ramos limpa o ar inspirado através da impactação de partículas em suspensão (CRAVEN
et al., 2007). O endoturbinados I, o mais dorsal e maior etmoturbinado, prolonga-se
rostralmente na CN, formando a base óssea da concha nasal dorsal; o endoturbinados II, ventral ao endoturbinados I, é a base da concha nasal média. A concha nasal ventral constitui uma formação própria do maxilar e não se considera como pertencente aos etmoturbinados, daí a sua denominação de maxiloturbinado (KÖNIG & LIEBICH, 2004).
Uma vez que o ar inalado abandona o vestíbulo nasal, desloca-se através dos meatos
nasais longitudinais, sendo eles o meato nasal dorsal (Meatus nasi dorsalis) (única via de
comunicação direta entre o vestíbulo nasal e o recesso olfativo) entre o teto de cada CN e a concha nasal dorsal, meato nasal médio (Meatus nasi medius) entre a concha nasal dorsal e a conha nasal ventral e o meato nasal ventral (Meatus nasi ventralis) localizado entre a concha nasal ventral e o teto do palato duro. O meato nasal comum (Meatus nasi communis) é um espaço longitudinal localizado em cada lado do septo nasal, ladeado lateralmente pelas conchas e sendo contínuo com os meatos nasais dorsal, ventral e médio. Finalmente, o meato
nasofaríngeo (Meatus nasopharyngeus) é o local onde os quatro meatos de cada CN
convergem e continua-se até às coanas, onde a nasofaringe se inicia (EVANS & LAHUNTA, 2013; BARRIOS et al., 2014).
7
A LC (Figura 3) é uma estrutura côncava, de protusão rostral, que se articula com o osso pré-esfenoide ventral e lateralmente e com os entalhes etmoidais do osso frontal, dorsalmente. Trata-se da verdadeira divisória anatómica entre a cavidade craniana e a CN. A lâmina apresenta-se perfurada por um grande número de forâmenes, na ordem dos trezentos e rondando 1,5 mm de diâmetro (EVANS & LAHUNTA, 2013), servindo como único local de passagem dos feixes dos neurónios sensoriais olfativos e fornecendo assim uma impressão óssea precisa da inervação olfativa periférica. As características morfológicas da LC, particularmente o seu tamanho, forma e número e distribuição dos seus forâmenes, variam entre diferentes mamíferos e entre diferentes animais da mesma espécie. Os tecidos que atravessam estes forâmenes incluem, além dos referidos feixes, apenas pequenas quantidades de tecido conjuntivo, líquido cefalorraquidiano e pequenos vasos sanguíneos, pelo que o número de forâmenes da LC é um bom indicador da quantidade de feixes nervosos que fazem a comunicação entre a mucosa olfativa e os BO. Assim a LC e os seus forâmenes pode ser usada como um indicador da performance olfativa do animal (BIRD et al., 2014). Estes forâmenes da LC (Foramina
laminae cribrosae) agrupam-se espacialmente segundo a disposição dos locais de fixação dos
turbinados. Um forâmen de maiores dimensões observado na superfície caudal da lâmina pode resultar da confluência de diversos pequenos forâmenes provenientes da superfície rostral. Estendendo-se craniomedialmente a partir dos bordos laterais da lâmina, pode observar-se uma elevação óssea livre de forâmenes, mas rodeada por forâmenes de maiores dimensões. Na porção mais caudal desta elevação, no local de comunicação do osso etmóide com o osso
pré-Figura 2: Corte sagital da CN do cão, evidenciando as conchas e meatos
8
esfenoide, encontram-se os forâmenes etmoidais (Foramina ethmoidalia), dois de cada lado. Estes dois forâmenes são o local de passagem das artérias e veias etmoidais, bem como o nervo etmoidal (EVANS & LAHUNTA, 2013). A superfície cranial da lâmina é bissectada ao longo da sua linha média pela lâmina perpendicular do osso etmoide, constituinte do septo nasal (BIRD et al., 2014). A crista galli divide a superfície caudal da LC na fossa direita e esquerda, denominadas fossas etmoidais da cavidade endocraniana (ROWE et al., 2005) que recebem cada uma delas os BO respetivos. Em cada lado da crista galli observa-se uma fila de forâmenes de grandes dimensões, sendo que um ou dois pares mais mediais destes forâmenes se abrem em tuneis mais profundos no osso que se continuam até à CN. Diversos pequenos forâmenes convergem numa direção cranial-caudal nestes tuneis de maiores dimensões. A localização destes grandes forâmenes na metade dorsal da lâmina é curiosa uma vez que se apresentam no local onde se pensa que os feixes de axónios provenientes do Órgão Vomeronasal (OV) atravessam desde o septo nasal até à cavidade craniana. Ainda não se sabe, no entanto, se algum forâmen da lâmina se dedica exclusivamente à passagem de nervos vomeronasais. O limite mais rostral destas fossas atinge o plano transversal que passa pelo centro das aberturas orbitais. A LC não se apresenta numa posição totalmente transversal: as metades esquerda e direita dispõe-se num plano quadispõe-se sagital e convergem rostralmente num angulo de aproximadamente 45 graus (EVANS & LAHUNTA, 2013).
Histologicamente, o revestimento interno da CN é feito por diferentes tipos de epitélio, sendo que o vestíbulo nasal, maxiloturbinado e etmoturbinados são revestidos por epitélio escamoso, respiratório e olfativo, respetivamente. A zona de transição do epitélio respiratório para olfativo (lâmina transversa) ocorre na região etmoidal cranial e é difícil de delimitar, podendo encontrar-se aqui aglomerações de células olfativas entre células não sensoriais
Figura 3: Secção transversal do crânio num plano
9
respiratórias. Nos animais macrosmáticos está descrita ainda a presença de um conjunto de espaços vasculares sob o epitélio, na porção ventral do septo nasal na região do maxiloturbinado que recebe a denominação de turbinado septal. A distensão ou contração desta estrutura tem a capacidade de regular o fluxo de ar inspirado consoante o seu estado (SANKARAN et al., 2012).
Com exceção do vestíbulo nasal mais rostral, toda a mucosa da CN é recoberta por secreções. Estas secreções aumentam as transferências de calor, ajudam a humidificar o ar inspirado, funcionam como barreira inicial contra químicos nocivos inspirados, absorvem as MO, auxiliam na remoção das partículas inspiradas através do transporte mucociliar e ainda garantem a sobrevivência dos cílios. A espessura desta camada de secreções varia entre os 5-30 mm e não é homogénea, sendo constituída pelo menos por duas fases e variando em composição química consoante sejam secreções da região respiratória ou olfativa (CRAVEN et
al., 2009).
A região olfativa da mucosa nasal (Regio olfactoria tunicae mucosae nasi) está localizada na porção dorsocaudal da CN incluindo alguma da superfície das conchas nasais do labirinto etmóide, meato nasal dorsal e septo nasal (EURELL & FRAPPIER, 2006) e continua-se, em extensão variável, com a parede lateral e medial da CN. Encontra-se recoberta por uma
mucosa olfativa especializada. Num tecido vivo, esta mucosa distingue-se pela sua coloração
(Figura 4) amarela acastanhada ou acinzentada (no caso do cão) devido aos pigmentos do
epitélio olfativo e das glândulas olfativas (Glandulae olfactoriae) associadas (ROSS, 2011)
bem como pela espessura do epitélio, maior que o da mucosa respiratória adjacente (EURELL & FRAPPIER, 2006). A área da mucosa nasal ocupada por epitélio olfativo varia grandemente entre espécies, ocupando entre 2 a 4 cm2 no homem, em torno de 18 cm2 no cão e 21 cm2 no gato (HILL et al., 2012). Em animais com o sentido do olfato mais desenvolvido, a área total ocupada pela mucosa olfativa é consideravelmente maior e em algumas raças de cães pode atingir mais de 150 cm2 (ROSS, 2011). A sensibilidade dos recetores olfativos individuais no cão não é necessariamente maior que a do homem, mas o facto de a região olfativa ser significativamente maior permite-lhe identificar MO em concentrações de 1 a 2 partes por trilião (CRAVEN et al., 2009) e 1:1000 vezes inferiores às identificadas pelo homem (REECE, 2009). O homem possui cerca de 107 recetores olfativos, enquanto o cão possui mais de 4-109 (HILL et al., 2012).
10
A lâmina própria (Lamina propria) ou córion da mucosa olfativa é contínua com o periósteo do osso subjacente (ROSS, 2011) e ao contrário do que acontece com o epitélio respiratório, não apresenta uma vascularização assinalável, não sofrendo por isso variações de espessura quando ocorre dilatação ou constrição vascular (CRAVEN et al., 2009). Este tecido conjuntivo contém diversos vasos sanguíneos e linfáticos, NO não mielinizados, nervos mielinizados e glândulas olfativas. O epitélio olfativo, tal como o epitélio respiratório, é pseudoestratificado colunar, mas apresenta diferentes tipos de células, não apresentando células de goblet como o epitélio respiratório (ROSS, 2011). O epitélio olfativo é constituído por
Células Neurosensoriais Olfativas, Células de Suporte, Células Basais e Glândulas Olfativas (Figura 5):
Células Neurosensoriais Olfativas ou Neurónios Olfativos – São neurónios bipolares que atravessam toda a espessura do epitélio e penetram no Sistema Nervoso Central (SNC) (ROSS, 2011). O epitélio olfativo possui milhares destes neurónios (LEDDO et al., 2005). A superfície apical de cada célula neurosensorial olfativa apresenta um prolongamento periférico ou processo dendrítico único e mais curto que se projeta até à superfície epitelial formando um abaulamento esférico denominado de vesícula olfativa. Um número variável de cílios finos (10 a 60) (CRAVEN et al., 2007) prolongam-se a partir da vesícula olfativa e estendem-se radialmente num plano paralelo à superfície epitelial, apresentando os recetores olfativos e sendo responsáveis por originar o impulso nervoso quando contactam com MO. Estes cílios de pequenas dimensões sobrepõem-se com os cílios das células neurosensoriais olfativas adjacentes, aumentando assim a área recetora. Os cílios são muito maiores que os presentes no epitélio respiratório e são descritos como não móveis. A membrana que reveste os cílios consiste numa bicamada fosfolipídica e a sua principal função é diferenciar os diferentes recetores olfativos, controlar as vias de fluxo de iões para dentro e para fora da célula, permitir
Figura 4: Vista lateral do septo nasal num cão adulto, mostrando a diferença de cor entre a mucosa olfativa
11
a deteção das MO e garantir a comunicação célula a célula (SANKARAN et al., 2012). A porção basal destas células corresponde a um processo axonal mais longo, fino e não mielinizado que abandona o compartimento epitelial, sendo dos mais pequenos axónios do sistema nervoso (HILL et al., 2012). Todos estes axónios seguem rodeadas pelas células de Schwann correspondentes (cada célula de Schwann envolve entre 10-100 axónios). Ao atravessarem a lâmina basal (Lamina basalis), os grupos de axónios provenientes das células neurosensoriais olfativas não formam um NO único, mas constituem um conjunto de feixes nervosos denominados de fila olfactoria (REECE et al., 2015) que atravessem a LC do osso etmóide bem como as meninges dura-máter e aracnoide e são envolvidos pela pia-máter, penetrando nos BO do telencéfalo (Telencephalon). O conjunto de todos estes feixes nervosos envolvidos por células da glia específicas, as ensheating cells (LEDDO et al., 2005) é que constitui o NO. As células sensoriais olfativas diferenciam-se a partir das células basais do epitélio, desenvolvendo as dendrites e axónios que lhe permitem comunicar com os neurónios de segunda ordem no BO (HILL et al., 2012). Diversos estudos mostram que o tempo de vida útil das células neurosensoriais olfativas é de aproximadamente um mês (30 a 60 dias) e que, em caso de lesão, são rapidamente substituídas por novas células derivadas das células basais (ROSS, 2011). Além disso apresentam uma outra característica única: à medida que a idade do animal aumenta, estas células têm períodos de vida cada vez maiores (SAKARAN et al., 2012).
Células de Suporte – São células colunares similares à neuróglia e que providenciam suporte mecânico e metabólico às células neurosensoriais olfativas. Têm a função de produzir e segregar proteínas de ligação à MO, as chamadas Proteínas de Ligação ao Odorante (PLO). As células de suporte são as mais numerosas do epitélio olfativo. Apresentam diversas microvilosidades na sua superfície apical, e um número elevado de mitocôndrias. Junções justaluminais podem ser identificadas entre estas células e as dendrites das células neurosensoriais olfativas (EURELL & FRAPPIER, 2006).
Células Basais – São as células-tronco do SO, a partir das quais as células de suporte e neurosensoriais olfativas se diferenciam. As células basais são pequenas e apresentam uma forma entre arredondado e poliédrico, localizando-se perto da lâmina basal. O seu citoplasma possui poucos organelos como ribossomas livres, uma característica coincidente com a sua função de células-tronco. As células basais caracterizam-se pela presença de complexos juncionais com outras células através de desmossomas (Figura 5) e de ligação à lâmina basal através de hemidesmossomas (EURELL & FRAPPIER, 2006). Outra característica que
12
justifica a sua função de diferenciação noutras células é a observação de processos de algumas destas células que embainham parcialmente a primeira porção dos axónios das células neurosensoriais olfativas, o que permite que mantenham uma relação com as células recetoras do olfato mesmo na sua forma indiferenciada (ROSS, 2011).
Glândulas olfativas ou Glândulas de Bowman - Características da mucosa olfativa, são glândulas serosas tubuloalveolares ramificadas que segregam as suas secreções proteicas através dos ductos para a superfície da mucosa olfativa. Grânulos de lipofuscina são frequentes nestas células, e em conjunto com os grânulos de lipofuscina das células de suporte, atribuem à mucosa a sua coloração característica. As secreções serosas da glândula olfativa funcionam como meio de dissolução para as MO. Um fluxo constante destas secreções desde a glândula até à superfície epitelial auxilia na eliminação contínua dos restos de MO já identificadas de forma a permitir reconhecimento de novas moléculas à medida que chegam ao epitélio (ROSS, 2011).
1.2.1.1.2 Nervo Olfativo (Par Craniano I)
Os NO ramificam-se na mucosa dos ectoturbinados, inervando aproximadamente metade dos etmoturbinados, a metade caudal do septo nasal e uma parte significativa do teto e paredes laterais da CN (Figura 6) (EVANS & LAHUNTA, 2013). Referidos normalmente e coletivamente como par craniano I, consistem na verdade num conjunto de numerosos axónios não mielinizados com corpos celulares localizados no epitélio olfativo da mucosa olfativa. Estes
Figura 5: Esquema representativo do epitélio olfativo: A, Células de
suporte; B, Células Basais; C, Axónios das células recetoras neurosensoriais; D, Dendrites; E, Porção fina distal dos cílios; F, Porção proximal grossa dos cílios; G, Complexos juncionais (Desmossomas) entre as células de suporte e as células neurosensoriais olfativas (adaptado de EURELL & FRAPPIER, 2006).
13
axónios nunca se ramificam e são notavelmente uniformes em diâmetro (0,2 a 0,4 µm). Após penetrarem na lâmina basal do epitélio olfativo, os segmentos proximais destes axónios migram através dos espaços vasculares extracelulares da fina camada submucosa das glândulas de Bowman. Uma vez que abandonam esta camada, inicialmente desorganizados e com espaços vazios entre eles, são envolvidos pelas células de revestimento olfativo (as ensheating cells), que condicionam uma compactação progressiva destas fibras nervosas, tornando-as alinhadas longitudinal e paralelamente para formar fascículos de máxima densidade. No NO como um todo, feixes individuais de células de revestimento olfativo estão sempre envolvidos pela sua própria lâmina basal e separados dos feixes vizinhos por um estreito espaço extracelular contendo algumas fibrilhas de colagénio. Estas células de revestimento são por sua vez rodeadas pelos chamados fibroblastos dos NO, extremamente longos e finos, estendendo-se em torno da circunferência das células de revestimento e separados das mesmas por um pequeno espaço contendo colagénio. Cada feixe de células revestimento pode ser envolvido por até seis processos de fibroblastos que os contornam. Dois processos de fibroblastos dos NO adjacentes estão em contacto direto entre si por complexos juncionais (FIELD et al., 2003).
Os axónios das células sensoriais olfativas penetram no crânio através da LC, na porção caudal da CN, para atingir os BO do telencéfalo, onde ocorre a sinapse (BARRET et al., 2010). Os axónios penetram no bulbo através dos diversos forâmenes da LC sobre a qual a superfície ventral do BO repousa. As células de revestimento acompanham os NO desde a mucosa olfativa, através da LC, até à camada nervosa do BO com a densidade de compactação e o alinhamento longitudinal a aumentar progressivamente e desaparecendo os espaços vazios entre os axónios, mas a envoltura de fibroblastos dos NO cessa a nível da camada mais externa do BO para permitir que as fibras nervosas de diferentes fascículos se misturem antes de penetrarem nos mesmos (FIELD et al., 2003).
Associados aos nervos olfativo encontra-se o nervo terminal (N. terminalis) e o nervo
vomeronasal (N. vomeronasalis), ambos provenientes do OV e que penetram respetivamente
no pedúnculo e BOA, formando um trato independente (BARRET et al., 2010). O nervo terminal é descrito como difícil de observar e a sua função é mal compreendida (EVANS & LAHUNTA, 2013).
14
O nervo trigémio (N. trigemini), par craniano V, não tem função olfativa direta mas é responsável por muitas dificuldades de determinação da origem de uma resposta sensorial perante estímulos externos. Este nervo e os seus ramos distribuem-se por toda a mucosa respiratória nasal e orofaríngea, incluindo uma ampla distribuição na mucosa olfativa e OV. As suas terminações nervosas podem ser excitadas por estímulos táteis, dolorosos e térmicos, mas também por odorantes. Embora possua menor sensibilidade para substâncias odoríferas que os recetores do SO, quase todos os odorantes em concentração suficiente podem levar à sua estimulação. Algumas perceções identificadas no animal durante o exame neurológico atribuídas ao olfato devem-se, na realidade, ao sentido trigéminal, uma vez que este intervém de forma total, parcial, direta ou indireta em todas as perceções identificadas através das funções gustativas e olfativas (BRITO, Fevereiro de 2010). As suas terminações livres localizadas na mucosa olfativa podem ser estimuladas por substâncias irritantes que originem determinados odores característico como seja pimenta, menta ou fumo de tabaco. A ativação destas fibras pode desencadear reações responsivas como espirros, lacrimejar, inibição respiratória entre outros reflexos (BARRET et al., 2010).
1.2.1.1.3 Rinencéfalo
O rinencéfalo apresenta várias funções ainda não totalmente esclarecidas, não apenas relacionado com o olfato. É constituído por uma porção basal relacionado com o olfato, uma porção septal que representa as áreas de associação dos reflexos de origem olfativa e uma porção límbica relacionada com as reações elementares provocadas por estímulos olfativos e os fenómenos vegetativos que acompanham as mesmas. É um termo adequado para designar o
Figura 6: Corte sagital da CN evidenciando a distribuição nervosa na mucosa septal e o OV (adaptado de EVANS
15
complexo do telencéfalo basal, hipocampo e estruturas associadas. Inclui entre outras os BO, tubérculos olfativos, tratos olfativos, núcleos olfativos anteriores, partes da amígdala e zonas do lobo piriforme (Nomina Anatómica Veterinária, 2012).
Os BO surgem na extremidade cranial de cada hemisfério cerebral (KAVOI & JAMEELA, 2011) dispostos a nível de cada uma das metades da LC e separados entre si pela
crista galli (ROWE et al., 2005). As células sensoriais olfativas projetam os seus axónios
através da LC como NO para o BO limitrofe ao SNC e que se encontra unido ao hemisfério cerebral corresponde através do pedunculo olfativo (Pedunculus olfactorius) (EVANS & LAHUNTA, 2013). Aqui contactam com as dendrites das células mitrais e células em tufo (cada axónio comunica com aproximadamente 15 dendrites) e interneurónios locais originando uma estrutura globular de unidades sinápticas denominada glomérulo olfativo (Figura 7) (BARRET et al., 2010), cada um com cerca de 50 a 200 µm de diâmetro num ratio de aproximadamente 1000:1 de axónios por BO. O cão apresenta cerca de 5000 glomérulos olfativos nos BO (HILDEBRAND & SHEPHERD, 1997) e cada glomérulo resulta da confluência de 5000 a 40000 axónios (LEDDO et al., 2005), apresentando assim elevados ratios de convergência aferente (HILDEBRAND & SHEPHERD, 1997).
As células em tufo são mais pequenas que as células mitrais e apresentam axónios mais finos mas ambas enviam os seus axónios para o córtex olfativo e parecem ser idênticas do ponto de vista funcional (SANKARAN et al., 2012). Além destes dois tipos de células, o BO apresentam ainda células periglomerulares (neurónios inibitórios que conectam os glomérulos entre si) e células granulares que não apresentam axónios mas realizam sinapses reciprocas com as dendrites laterais das células mitrais e em tufo. Nestas sinapses as células mitrais excitam as células granulares libertando glutamato, enquanto as células granulares inibem as primeiras libertando GABA (BARRET et al., 2010). Devido à proximidade dos axónios no percurso e penetração nos glomérulos olfativos, estabelecem-se ainda entre estes transmissões efáticas que permitem sincronizar potenciais de ação com as fibras vizinhas com precisão de milissegundos a partir do campo elétrico extracelular (LEDDO et al., 2005).
16
Do ponto de vista histológico, sete camadas constituem a parede dos BO. Da camada mais superficial para a mais profunda apresentam-se (Figura 7): (1) camada de nervos
olfativos (constituída por axónios dos NO e células da glia), (2) camada glomerular (vários
axónios olfativos em sinapse com células mitrais, em tufo e interneurónios dentro de glomérulos olfativos), (3) camada plexiforme externa (onde se encontram os núcleos das células em tufo, dendrites secundárias das células mitrais e em tufo bem como as dendrites ramificadas das células granulares), (4) camada de células mitrais (corpos celular cónicos de células mitrais alinhados numa só fileira e alguns núcleos de células granulares), (5) camada plexiforme
interna (mais fina que a camada plexiforme externa, sem corpos celulares e contendo apenas
os axónios das células mitrais e em tufo), (6) camada granular (com a maior parte dos núcleos
Figura 7: Diagrama esquemático representando as camadas histológicas do BO, as interações entre as diferentes
células bulbares e os circuitos neurais básicos no BO. Todos os axónios das células sensoriais olfativas se projetam para determinado glomérulo olfativo (GO) consoante o tipo de recetor que apresentam; PG, Células Periglomerulares; T, Células em Tufo; M, Células Mitrais; G, Células Granulares (adaptado de DUDA, 2009).
17
das células granulares) e (7) camada subependimária (constituída por neurónios a entrar e abandonar o BO) (DUDA, 2009; EVANS & LAHUNTA, 2013;). Um número substancial de fibras centrífugas penetra no bulbo através da camada subependimária, em direção à camada granular e glomerular, as duas onde existe maior número de neurónios inibitórios (LEDDO et
al., 2005).
Os principais outputs dos BO são as células mitrais que se localizam na camada de células mitrais. A dendrite primária (ou apical) de cada célula mitral estende-se verticalmente a partir do corpo celular e comunica com um único glomérulo onde ocorrem interações massivas com interneurónios bulbares que se localizam nas terminações dos axónios olfativo. Em contraste com a dendrite primária, a dendrite secundária (ou basal) estende-se horizontalmente de forma radial, cerca de 1,000 µm, para abranger a quase totalidade do BO. Na camada plexiforme externa, interagem com interneurónios inibitórios denominados de células granulares, a população de células mais numerosa do BO (DUDA, 2009). Tanto os
inputs excitatórios como os circuitos intrabulbares, que basicamente incluem duas ligações
distintas entre dendrites primárias e células periglomerulares e entre dendrites secundárias e células granulares, controlam rigorosamente a informação que abandona o BO. A grande diferença entre as células periglomerulares e granulares é que as primeiras medeiam maioritariamente as interações entre células afiliadas com o mesmo glomérulo enquanto as células granulares vão mediar as interações entre neurónios que abandonam diferentes glomérulos. Muitos neurónios bulbares comunicam entre si via sinapses dendrodentríticas recíprocas. Assim a propagação dos potenciais de ação e da mensagem excitatória através do BO para as vias olfativas seguintes é controlada por diversos mecanismos, garantindo uma restrição na informação que segue para os níveis sequentes (LEDDO et al., 2005).
O pedúnculo olfativo (Figura 8) inclui o núcleo olfativo anterior e prolonga-se desde o BO até ao tubérculo olfativo (Tuberculum olfactorium) que ocupa uma posição medial, estando o córtex piriforme lateralmente na porção mais caudal (BRUNJES et al., 2005). O pedúnculo olfativo está separado do lobo frontal pelos sulcos rinal lateral (Sulcus rhinalis
lateralis) e medial (Sulcus rhinalis medialis) que se encontram ligeiramente dorsais a este. Na
sua superfície ventrolateral observa-se facilmente o trato olfativo lateral (Tractus olfactorii
lateralis) que começa como uma espessa banda a nível do BO que coalesce num trato bem
visível e diferenciável (EVANS & LAHUNTA, 2013). O núcleo olfativo anterior é responsável por regular a informação que provém do BO e segue para o córtex piriforme, bem como por
18
regular o circuito contrário. Além disso também permite a troca de informação entre os BO direito e esquerdo bem como para os córtex olfativo direito e esquerdo. A porção bulbar da comissura rostral transporta as chamadas fibras centrífugas, axónios provenientes do córtex olfativo e que se dirigem para os glomérulos olfativos dos BO, conseguindo modular a resposta dos mesmos à chegada de informação olfativa (BRUNJES et al., 2005).
O tubérculo olfativo apresenta-se como uma protuberância ventral, caudal ao pedúnculo olfativo e rostral ao giro diagonal (Gyrus diagonalis). É constituído por um córtex de três camadas que envia axónios para o hipotálamo (Hypothalamus) através do feixe medial do
prosencéfalo ou fascículo medial do telencéfalo (Fasciculus medialis telencephali) (EVANS
& LAHUNTA, 2013).
O lobo piriforme (Lobus piriformis) está relacionado com a olfação consciente. Recebe axónios das células mitrais do BO através do trato olfativo lateral. Consiste numa estrutura aplanada demarcada da proeminência caudal através da fossa lateral cerebral (Fossa lateralis
cerebri) e a porção mais rostral é demarcada pelo tubérculo olfativo. Inclui o giro olfativo
lateral (Gyrus olfactorius lateralis), que faz parte do córtex prépiriforme. A porção mais caudal
do lobo piriforme é proeminente devido à presença da amígdala (Amygdala), hipocampo
(Hippocampus) e ventrículo lateral (Ventriculus lateralis)em profundidade. A proeminência é recoberta por diferentes tipos de córtex. O lobo piriforme continua-se caudalmente com o giro
do parahipocampo (Gyrus parahippocampalis) (EVANS & LAHUNTA, 2013).
Figura 8: Vista ventral do cérebro: 1, BO; 2,
Tubérculo Olfativo; 3, Giro diagonal; 4, Comissura Rostral; 5, Colunas do Fórnix; 6, Cápsula Interna; 7, Corpo do Fórnix; 8, Giro do Parahipocampo; 9, Sulco do Hipocampo; 10, Giro Caloso; 11, Flexura Subsplenial do Giro Dentado; 12, Tubérculo do Giro Dentado; 13, Lobo Occipital; 14, Giro Dentado; 15, Ramo do Sulco Rinal Lateral; 16, Porção Caudal do Sulco Rinal Lateral; 17, Porção Intermédia do Sulco Rinal Lateral; 18, Lobo Piriforme; 19, Porção Rostral do Sulco Rinal Lateral; 20, Pedúnculo Olfativo; 21, Sulco Rinal Medial; (adaptado de EVANS & LAHUNTA, 2013).
19
A amígdala ou Corpo Amigdalóide (Corpus amygdaloideum), assim denominado devido à sua forma de amêndoa, localiza-se no lobo piriforme. O chamado complexo amigdaloide possui aproximadamente 12 núcleos, incluindo seis de maiores dimensões agrupados em três grupos. A amígdala está relacionada com reações emocionais, principalmente aquelas relacionadas com informação sensitiva. É um componente maior do
sistema límbico. (EVANS & LAHUNTA, 2013).
A região septal localiza-se rostralmente à comissura rostral e ventral ao joelho e rostro do corpo caloso (Corpus callosum). O giro paraterminal (Gyrus paraterminalis) constitui a superfície medial da parede septal e núcleos septais (Nuclei septi) e está incluído nesta parede. Estes núcleos recebem aferências axonais do trato olfativo medial (Tractus olfactorii medialis) e apresentam comunicação recíproca com a amígdala [através da estria terminal (Stria
terminalis)], hipocampo [através do fórnix (Fornix)], hipotálamo (através do feixe medial do
prosencéfalo) e habénula (Habenula) [através da estria habenular (Stria habenularis)]. A
porção rostral (Pars rostralis) ou bulbarda comissura rostral surge rostralmente ao longo da face ventral da capsula interna e penetra no pedúnculo olfativo, originando o trato olfativo
intermédio (Tractus olfactorius intermedius). Estes axónios comunicam de forma recíproca
com os BO e os núcleos olfativos anteriores de cada lado, terminando na camada granular do BO (EVANS & LAHUNTA, 2013).
1.2.1.2 Sistema Olfativo Acessório
O OV, também conhecido como órgão de Jacobson, é um órgão par localizado rostralmente à base do septo nasal, ventralmente ao epitélio olfativo principal (HILL et al., 2012) (Figura 4 e Figura 6), tratando-se de um órgão tubular em fundo de saco cego, revestido internamente por epitélio [ducto epitelial vomeronasal (Ductus vomeronasalis)], uma submucosa intermédia e parcialmente revestido externamente por um suporte de cartilagem
vomeronasal (Cartilago vomeronasalis) (EURELL & FRAPPIER, 2006; EVANS &
LAHUNTA, 2013). Cada OV tem aproximadamente 80 mm de comprimento e abre-se cranialmente num ducto incisivo que comunica a CN com a cavidade oral. Na cavidade oral o ducto incisivo abre-se a nível da estrutura denominada papila incisiva (Papilla incisiva), entre os dois dentes incisivos, enquanto na CN a abertura do ducto é difusa, podendo ser considerada como uma projeção da mucosa que cobre os bordos da fissura palatina (Fissura palatina). O ducto apresenta uma forma de lua crescente em corte transversal, com a mucosa lateral convexa