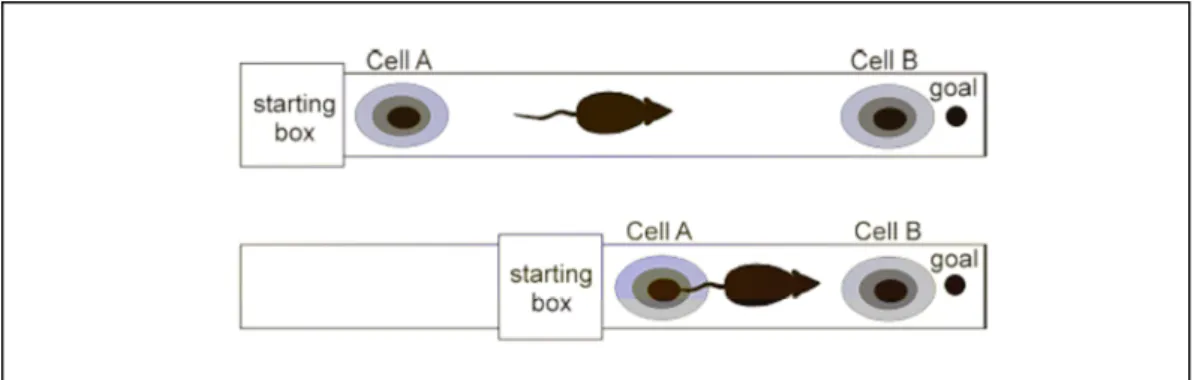
Neuroanatomie de la mémoire spatiale
Introduction
En 1978, O'Keefe et Nadel émettent l'hypothèse que l'hippocampe constitue le substrat neuroanatomique des représentations spatiales allocentriques (cartes cognitives). Cette hypothèse reposait sur des données lésionnelles et était particulièrement étayée par la découverte de cellules hippocampiques.
La région hippocampique
- Définition
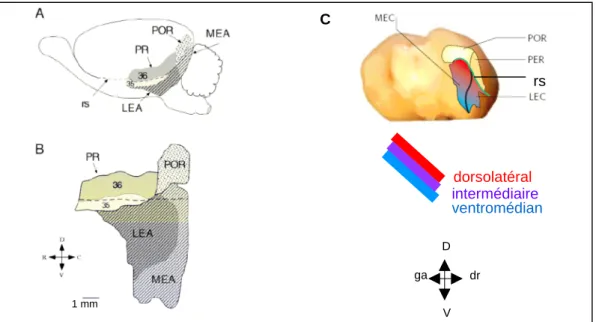
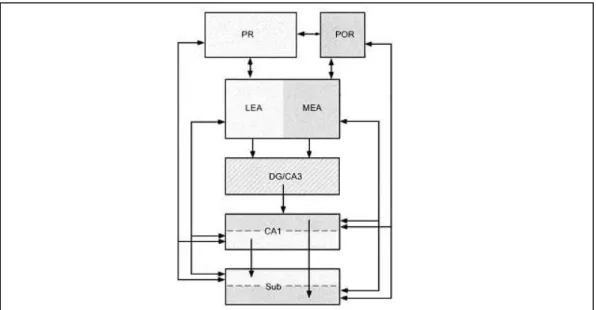
- La formation hippocampique
- Région parahippocampique
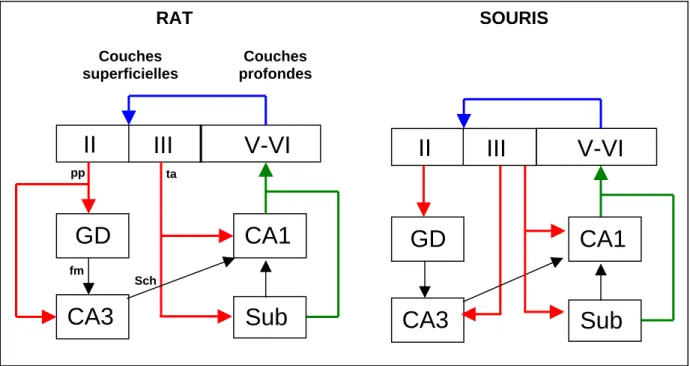
- Différences rat / souris concernant la région hippocampique
Le cortex entorhinal est situé à l'interface de l'hippocampe et du cortex parahippocampique (Burwell, 2000 ; Witter et al., 1989 ; Lavenex et Amaral, 2000). Le cortex entorhinal médial (MEC) reçoit principalement des projections du POR, tandis que le cortex entorhinal latéral (LEC) reçoit davantage du PER.
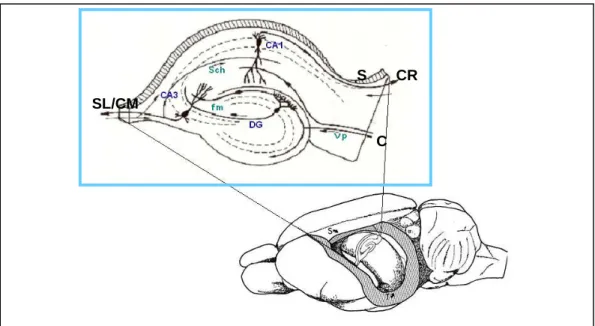
Autres régions corticales impliquées dans la mémoire spatiale
- Cortex pariétal
- Cortex préfrontal
De plus, l’activité de ces cellules est indépendante de la direction de la tête de l’animal. La comparaison des paramètres de décharge cellulaire entre les deux groupes (zif268 et contrôle) dans S1 ne révèle aucune détérioration chez les souris mutantes (voir l'article 2 ci-dessous pour plus de détails).
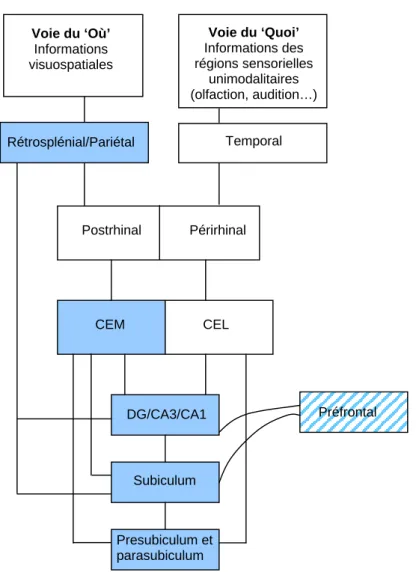
Les bases cellulaires de la mémoire spatiale
Introduction
Cependant, l’argument essentiel en faveur de la fonction spatiale de l’hippocampe repose sur l’existence de « cellules de lieu ». Dans ce chapitre, je passerai donc en revue les principales propriétés de décharge ainsi que les déterminants sensoriels de l'activité des cellules de ville, des cellules de direction et des cellules de grille.
Cellules de lieu
- Propriétés
- Distribution et forme des champs d’activité
- Contrôle sensoriel de l’activité des cellules de lieu
Enfin, au niveau de la population neuronale, l'activité des cellules de lieu serait la traduction électrophysiologique de la représentation des positions de l'animal. Les informations tactiles sont également importantes pour contrôler l’activité des cellules localisées.
Cellules de direction de la tête
- Propriétés et points communs avec les cellules de lieu
- Contrôle sensoriel de l’activité des cellules de direction
- Régions anatomiques qui possèdent des cellules de direction
Cartes de débit montrant une structure triangulaire identique (espacement, taille du sous-champ et fréquence de décharge du réseau), quelle que soit la taille de l'environnement. Ainsi les cellules générales (14 cellules + 6 cellules) ont des champs contrôlés par les index restants (non contrôlés et contrôlés) lors de la session CR.
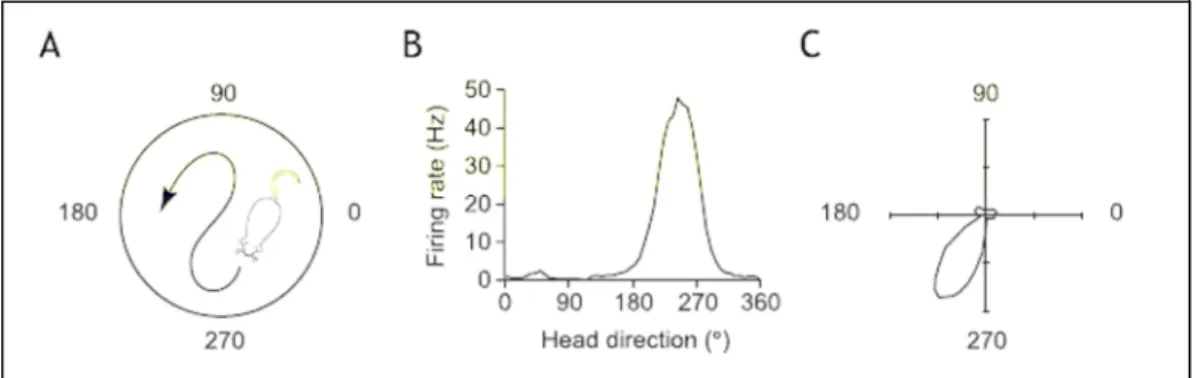
Relations entre cellules de lieu et les cellules de direction
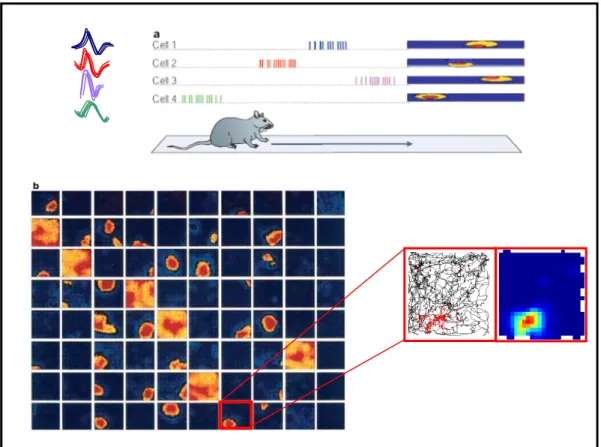
Cellules grille
- Origines de la découverte
- Propriétés
- Influence des informations sensorielles sur l’activité des cellules grille
L'enregistrement dans un environnement très vaste (2 m de diamètre) révèle le motif très régulier caractéristique de l'activité spatiale d'une cellule de grille dans la couche superficielle (II) du CEM. L’une des propriétés fondamentales des cellules de la grille est que leur schéma d’activité ne dépend pas de la nature de l’environnement dans lequel se trouve l’animal.
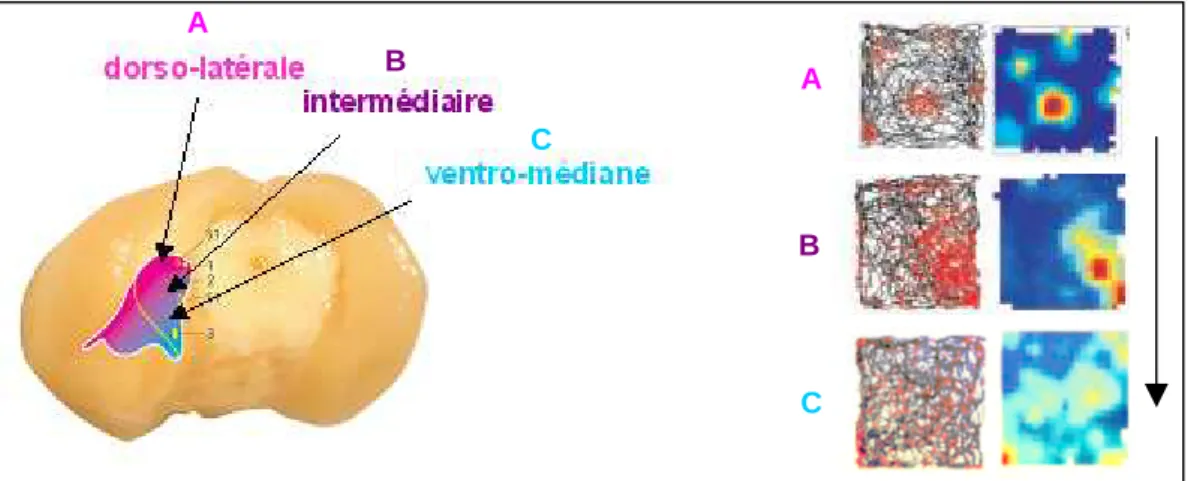
Autres cellules de la formation hippocampique : cellules du complexe subiculaire
- Subiculum
- Pré- et para-subiculum
L'effet d'un changement d'environnement sur les champs d'activité des cellules du céciculum et de l'hippocampe. Lors du passage d'un environnement rond à un environnement carré, la plupart des champs cellulaires de l'hippocampe présentent un remappage, c'est-à-dire créer des représentations distinctes, tandis que les champs d'activité des cellules du subiculum restent stables.
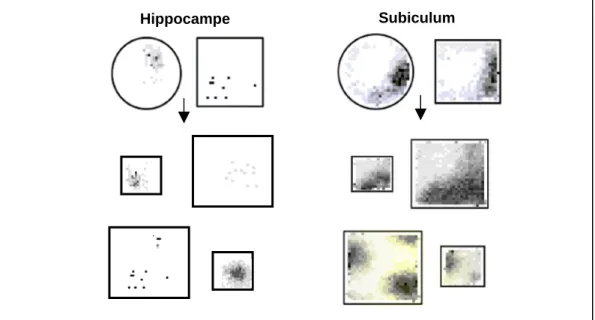
Conclusion sur les cellules avec des corrélats spatiaux
L’hippocampe: mémoire de l’espace ou espace de mémoire ?
La mémoire à long terme de la représentation spatiale d'un nouvel environnement est fortement altérée chez les souris mutantes pour le gène zif268. La formation d’une nouvelle représentation semble donc interférer avec la réactivation de la représentation de l’environnement familier.

Les mécanismes fondamentaux de la flexibilité de la représentation spatiale
Description du phénomène
On sait que l'organisation spatiale des champs d'activité d'une population de cellules de lieu est stable dans un environnement donné. Dans les deux études, les expérimentateurs ont déplacé les animaux d’un environnement (rond) à un autre environnement (carré).
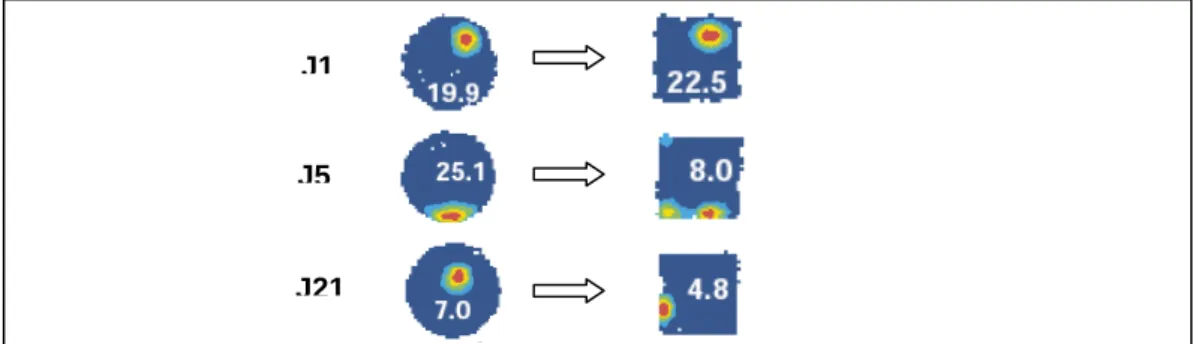
Les différents types de remapping décrits dans la littérature
Cette recartographie implique un changement imprévisible de localisation et un changement dans la fréquence des activités sur le terrain à mesure que l'animal se déplace d'un environnement à un autre. Lors de l'enregistrement de plusieurs cellules en même temps, des transformations de l'environnement peuvent provoquer une remapping (disparition, apparition, changement de localisation) des champs situés à proximité de la modification.
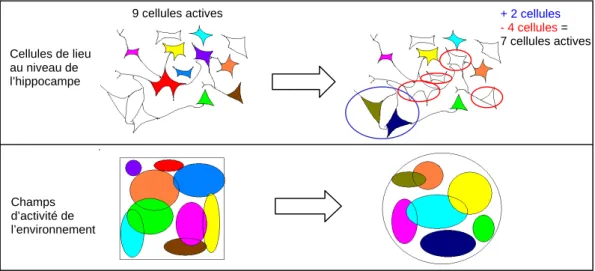
Pattern separation/completion et adaptation des cellules de lieu aux modifications
- Théorie des attracteurs
- Différence CA1/CA3
- Gyrus denté
- Mémoire des représentations : importance d’un comparateur
Ainsi, schématiquement, CA3 répondrait de manière non linéaire aux changements de l'environnement, tandis que CA1 répondrait de manière linéaire (Guzowski et al., 2004) (Figure 3.8). Cette représentation serait comparée à la représentation des expériences antérieures enregistrées dans CA3 (Hasselmo et al., 1995).
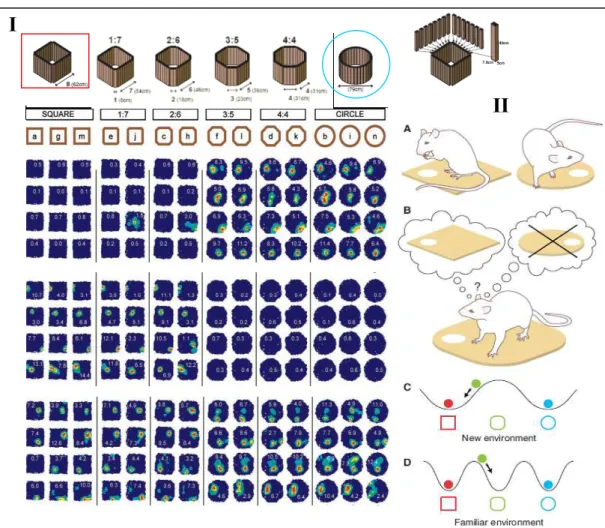
Cellules grille et « remapping » hippocampique
Si l'on suppose que les représentations spatiales sont stockées au niveau du néocortex, il serait plausible d'y trouver des cellules dont l'activité est liée à la position de l'animal dans l'environnement. Notons que le transfert d'informations peut être facilité par les phénomènes de réactivation des représentations spatiales pendant le sommeil (Wilson et McNaughton, 1994).
Conclusion
L'injection d'un inhibiteur de la protéine kinase C dans l'hippocampe entraîne des déficits de mémoire spatiale à long terme (Pastalkova et al., 2006). La reconsolidation de la mémoire de reconnaissance d'objets est altérée chez les souris mutantes zif268-/-.
Les mécanismes moléculaires de la flexibilité de la représentation spatiale
Cellules de lieu et plasticité synaptique
- La potentialisation à long terme (PLT) à ses origines
- La PLT, un modèle de plasticité impliqué dans les processus mnésiques
- Y-a-t-il un lien entre la PLT et la mémoire spatiale?
Certaines études ont testé l'implication des récepteurs NMDA (N-méthyl-D-aspartate) - connus pour être importants dans l'induction de la LTP - dans l'apprentissage. En effet, les protocoles d’induction de LTP sont par définition basés sur des stimulations artificielles et non physiologiques.
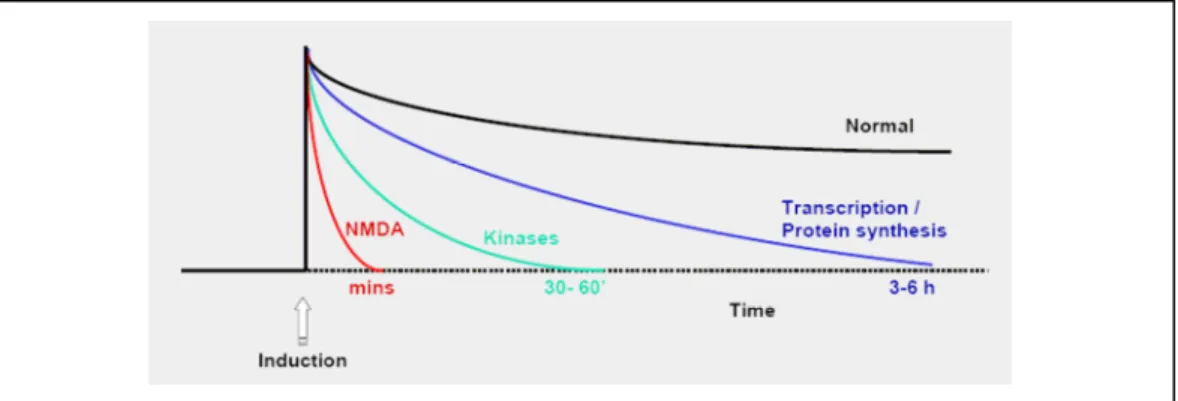
Zif268 et mémoire
- Qui est zif268 ?
- Zif268 et la consolidation des souvenirs
- Zif268 et la re-consolidation des souvenirs
Cela suggère que zif268 est un gène impliqué dans la reconsolidation de la mémoire objet. Pour résumer, le gène zif268 apparaît donc nécessaire à la fois à la consolidation et à la reconsolidation de la mémoire de reconnaissance (Jones et al., 2001 ; Bozon et al., 2003).
Zif268 et cellules de lieu ?
Puis, dans une deuxième partie (article n°2), j'ai étudié les aspects mémoriels de la flexibilité de ces représentations spatiales, grâce à l'enregistrement de l'activité des cellules de la ville. L'angle correspondant à RMax permet d'évaluer la valeur de rotation de la zone d'activité d'une séance à la suivante.
Matériels et méthodes communs
Animaux
- Rats
- Souris
Enfin, le troisième type de manipulation génétique consiste en un contrôle temporel du début de l'expression du gène d'intérêt (knockouts inductibles). De plus, l'expression du gène LacZ constitutif et induite par le PLT est comparable à l'expression du gène zif268 chez les souris témoins, ce qui indique que la cascade de signalisation se situe en amont.
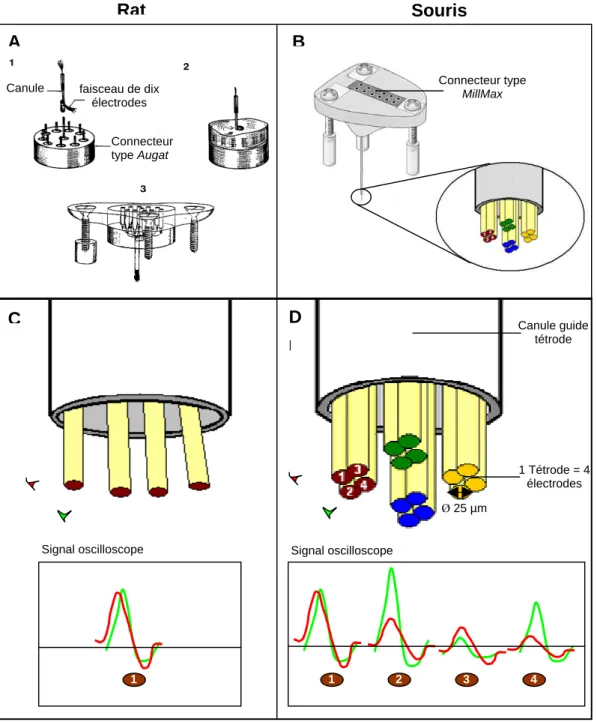
Fabrication des plots porte-électrodes
- Rat
- Souris : tétrodes
La séparation des signaux est basée sur l'amplitude relative des potentiels d'action qui apparaissent simultanément sur les 4 électrodes de la tétrode. Dans l'expérience sur le rat (article 1), j'ai utilisé 10 électrodes monofilaires, tandis que dans l'expérience sur la souris (article 2), j'ai utilisé quatre tétrodes (16 électrodes).
Entraînement pré-chirurgie
- Rat
- Souris
Chirurgie
- Rat
- Souris
Une fois le tenon en place, le trou d'implantation est recouvert de vaseline stérile et de ciment dentaire pour fixer les manchons aux vis ancrées dans le crâne de l'animal. La pointe des tétrodes est placée sur l'hippocampe droit aux coordonnées suivantes : AP = -2 mm, L = -2 mm par rapport au bregma, DV = -0,7 mm par rapport à la surface du cerveau.
Technique d’enregistrement
- Pièce d’enregistrement
- Enregistrement
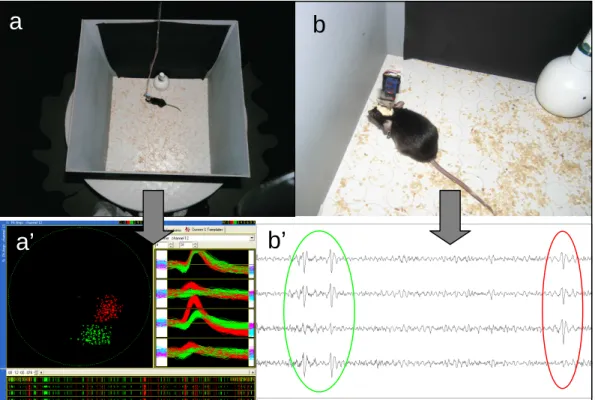
Le but de la technique utilisée est d'associer chaque événement électrophysiologique à la position prise par l'animal au moment où l'événement s'est produit. Un signal de position donné par la LED sur la tête de l'animal est capté par une caméra qui enregistre les coordonnées du rat.
Analyse des données
Une carte de fréquence d’activité est construite à partir de la combinaison de ces deux informations. . étant donné, l'élément correspondant de la matrice est augmenté d'une unité). Le programme calcule ensuite la fréquence d'activité moyenne en divisant la première matrice par la seconde, pixel par pixel.
Caractéristiques numériques
Un champ d'activité est défini comme étant constitué d'au moins 9 pixels contigus dans lesquels la fréquence d'activité est supérieure à la fréquence moyenne calculée sur l'ensemble de la session. Nous pouvons évaluer quantitativement la stabilité du terrain entre deux sessions en fonction de la corrélation spatiale entre les deux cartes d'activité.
Développement d’un nouveau système d’enregistrement de l’activité neuronale
Ceci suggère que le système de cellules de lieu peut montrer des réponses différentes selon la nature des signaux utilisés. Il n'y a aucune différence dans la stabilité à court terme de la représentation dans le nouvel environnement (S3/S5) entre les souris zif268 et les souris témoins.
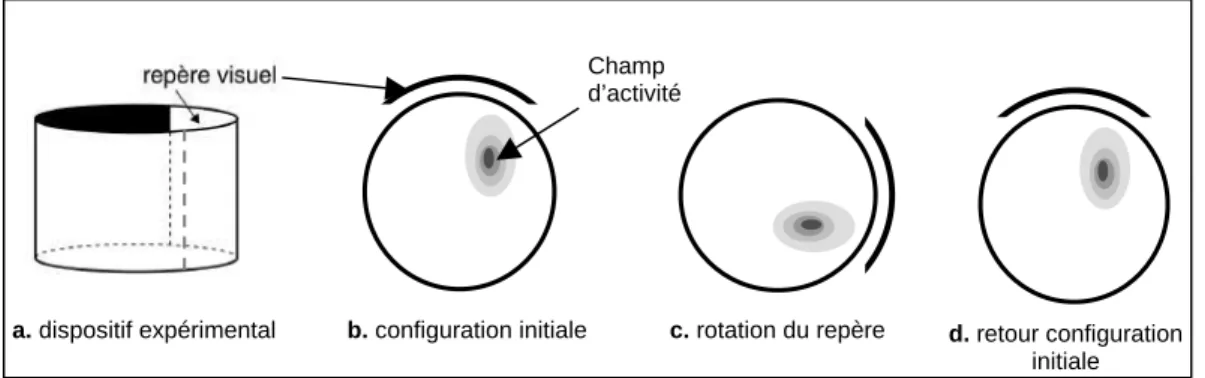
Flexibilité des cellules de lieu dans l’utilisation des repères proximaux et distaux
Introduction
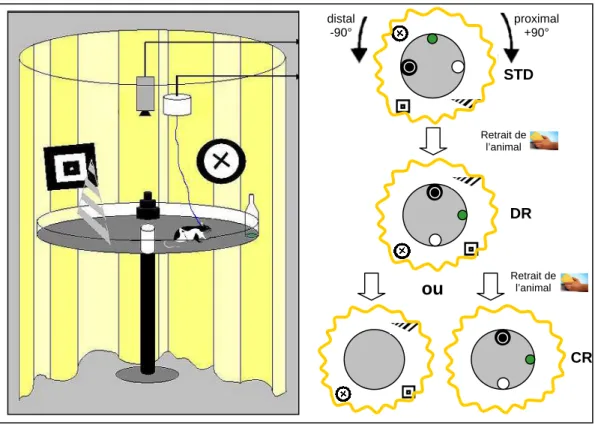
Nous avons donc enregistré l'activité des cellules de lieu chez le rat dans une arène circulaire, contenant trois objets, dits proximaux, placés à la périphérie de l'arène. Ensuite, nous avons procédé à la suppression de l'un des deux ensembles d'indices (proximaux ou distaux) pour déterminer si les cellules de lieu peuvent s'adapter de manière flexible à la dégradation de l'environnement.
Matériels et méthodes
- Animaux
- Dispositif expérimental
- Méthodes d’enregistrement
- Protocole expérimental
Parfois, une séance de retrait (RC) d'objets proximaux ou de signes distaux est réalisée. Les rats n'ont jamais été exposés à une double rotation ou à la suppression de signaux avant l'enregistrement cellulaire.
Analyse des données
Résultats
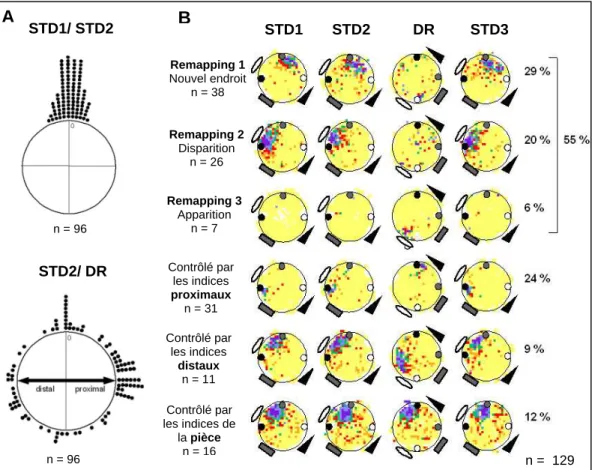
Certaines cellules sont contrôlées par des objets proximaux, d'autres par des signaux distaux et d'autres encore présentent une recartographie. L'examen visuel des cartes d'activité des 12 cellules comportant un champ CR suggère que les champs d'activité sont contrôlés par les signaux restants.
Discussion
Nous avons constaté, comme Shapiro et al., que la majorité des cellules de la ville étaient contrôlées par la configuration des personnages (55 %). Une possibilité est que les cellules réagissent individuellement aux signaux environnementaux (Shapiro et al., 1997 ; Knierim, 2002).
Introduction
Le gène précoce immédiat (IEG) zif268 est nécessaire à la stabilité à long terme des cellules de place CA1 de l'hippocampe. Parce que Zif268 est un élément essentiel au maintien de la LTP dans le temps, il devrait également être nécessaire à la stabilité à long terme des cellules de lieu et donc à la stabilité de la mémoire spatiale.
Matériels et méthodes
- Animaux
- Dispositif expérimental et méthodes d’enregistrement
- Protocole expérimental
Un objet (bouteille en plastique blanche) est placé devant cette carte pour renforcer l'identité du milieu (figure 7.1). Lorsqu’une cellule est localisée, après deux premières sessions d’enregistrement dans l’environnement carré, les souris sont exposées à un nouvel environnement.

Analyse des données
Les séances 1 à 5 ont lieu le premier jour, tandis que les séances 6 et 7 ont lieu 24 heures après la fin de la dernière séance du premier jour. Nous nous posons les questions suivantes : 1) Entre S1 et S2, les champs d'activité sont-ils stables à conditions constantes ? 2) Les cellules de S3 sont-elles capables de former une nouvelle représentation ? 3) Les cellules de lieu en S5 peuvent-elles réactiver la nouvelle représentation après un court délai (1 heure) ? 4) Les cellules placées dans S7 peuvent-elles réactiver la nouvelle représentation après un long délai (24 heures). Entre les séances, les souris sont remises dans leurs cages et le sol de l'arène est nettoyé pour neutraliser les signaux olfactifs.
Résultats
Formation et stabilité de champs d'activité dans l'environnement familier (entre S1 et S2) comparables chez les souris zif268-/- et chez les souris témoins. Exemples de cartes d'activité représentatives de l'instabilité (remappage rotationnel) des champs d'activité à court terme dans l'environnement familier (S2/S4) après la séance dans le nouvel environnement (cercle) chez des souris zif268.
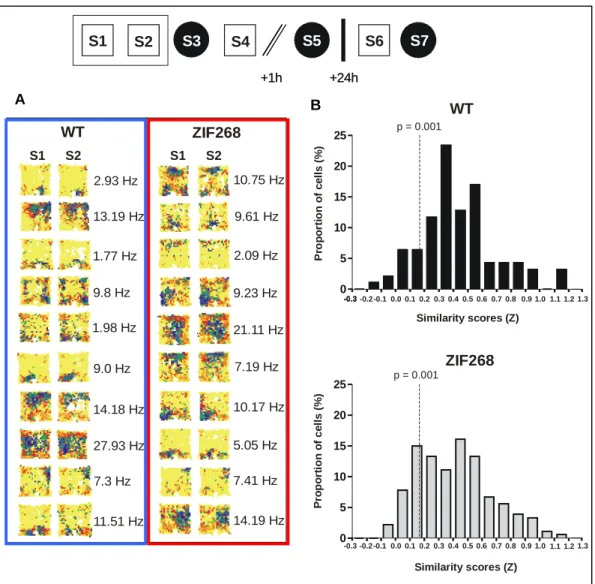
Discussion
Par exemple, les souris présentant une mutation CaMKII présentent un déficit en PLT et. Par conséquent, il est possible que zif268 soit important pour la stabilisation à long terme de la représentation des cellules de lieu, même après une exposition environnementale répétée.
Le gène zif268 est nécessaire à la re-consolidation de la représentation spatiale
Introduction
Ce processus était appelé reconsolidation. 2002) ont déjà montré que les souris mutantes zif268, dont les performances mnésiques ont été évaluées lors d'une tâche de détection de changement spatial 24 h après l'apprentissage, présentaient des déficits en consolidation de la mémoire spatiale. Dans cette étude, nous étudions si zif268 est impliqué dans les processus de reconsolidation de la mémoire spatiale.
Matériels et méthodes
- Animaux
- Tâche et protocole expérimentaux
Une autre étude a également montré que les souris mutantes zif268 présentaient des déficits de reconsolidation dans une tâche impliquant la mémoire de reconnaissance d'objets (Bozon et al., 2003). Cette étude nous permettra également de mieux comprendre les déficits de stabilité des représentations spatiales qui surviennent spécifiquement chez les souris zif268 dans l'environnement.
Analyse des données
Résultats
Discussion