Journal of Environmental Management
Texto
Imagem
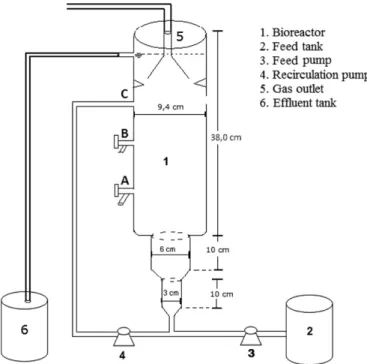
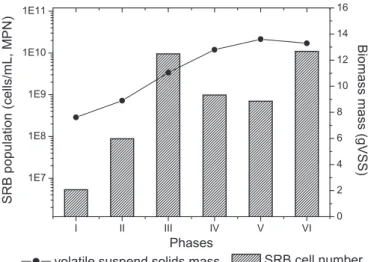
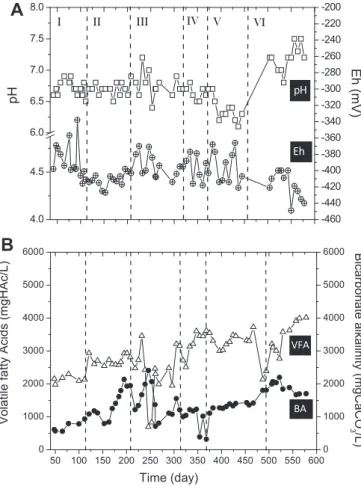

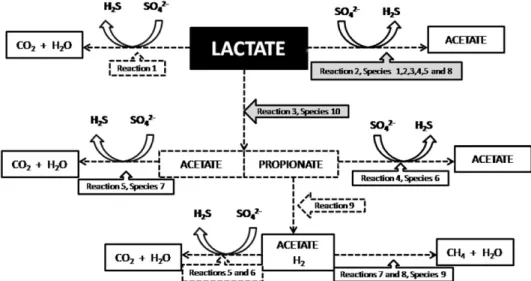

Documentos relacionados
Conclusion: The baseline blood lactate of SS and SC individuals is 3 to 4 times higher than that of healthy subjects, probably due to the fact that these patients have a
However, the blood lactate observed during swimming against water current suggests that dourado is not a lactate releaser, in spite of the high values reached during the
Starting with L-alanine and L-lactate that entered in the gluconeogenic pathway at pyruvate step, the present results suggested that the treatment with glucose
The reaction of isatin and 1-alkylisatins with primary alkylamines yields the corresponding 3-imines, which upon reduction with sodium borohydride in hot ethanol
When the possible transition structures for the reaction of the dicoordinated complexes of piperonal with AlCl 3 were calculated (Scheme 3), the barrier of activation for the reaction
These results suggest that the cathodic peak current i pc (II) results from the reduction of a product formed by the reaction between citrate and some product formed during DA
Concerning DO3 and DB3, the main oxidation product was 4,4’-dinitroazobenzene, originated from the oxidation of the terminal amine in the same way that the reaction occurred in
The results obtained related to the plasmatic levels of lactate, which permit the indirect evaluation of the metabolic acidosis during the exercise, demonstrated that the
