h ttp : / / w w w . b j m i c r o b i o l . c o m . b r /
Environmental
Microbiology
Anaerobic
biodegradation
of
benzo(a)pyrene
by
a
novel
Cellulosimicrobium
cellulans
CWS2
isolated
from
polycyclic
aromatic
hydrocarbon-contaminated
soil
Wei
Qin
a,
Fuqiang
Fan
b,
Yi
Zhu
a,
Xiaolong
Huang
a,
Aizhong
Ding
a,
Xiang
Liu
c,
Junfeng
Dou
a,∗aBeijingNormalUniversity,CollegeofWaterSciences,Beijing,China
bMemorialUniversityofNewfoundland,FacultyofEngineeringandAppliedScience,NorthernRegionPersistentOrganicPollution
ControlLaboratory,St.John’s,Canada
cTsinghuaUniversity,SchoolofEnvironment,Beijing,China
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received30December2016 Accepted25April2017
Availableonline12October2017
AssociateEditor:LucySeldin
Keywords: Anaerobic Biodegradation Benzo(a)pyrene
CellulosimicrobiumcellulansCWS2
a
b
s
t
r
a
c
t
CellulosimicrobiumcellulansCWS2,anovelstraincapableofutilizingbenzo(a)pyrene(BaP)as thesolecarbonandenergysourceundernitrate-reducingconditions,wasisolatedfrom PAH-contaminatedsoil.TemperatureandpHsignificantlyaffectedBaPbiodegradation,andthe strainexhibitedenhancedbiodegradationabilityattemperaturesabove30◦Candbetween
pH7and10.ThehighestBaPremovalrate(78.8%)wasobservedin13dayswhentheinitial BaPconcentrationwas10mg/L,andthestraindegradedBaPatconstantrateevenatahigher concentration(50mg/L).MetalexposureexperimentalresultsillustratedthatCd(II)wasthe onlymetalionthatsignificantlyinhibitedbiodegradationofBaP.Theadditionof0.5and 1.0g/LglucoseenhancedBaPbiodegradation,whiletheadditionoflow-molecular-weight organicacidswithstrongeracidityreducedBaPremovalratesduringco-metabolic biodegra-dation.Theadditionofphenanthreneandpyrene,whichweredegradedtosomeextent bythestrain,showednodistincteffectonBaPbiodegradation.Gaschromatography–mass spectrometry(GC-MS)analysisrevealedthatthefiveringsofBaPopened,producing com-poundswithonetofourringswhichweremorebioavailable.Thus,thestrainexhibited strongBaPdegradationcapabilityandhasgreatpotentialintheremediationof BaP-/PAH-contaminatedenvironments.
©2017SociedadeBrasileiradeMicrobiologia.PublishedbyElsevierEditoraLtda.Thisis anopenaccessarticleundertheCCBY-NC-NDlicense(http://creativecommons.org/ licenses/by-nc-nd/4.0/).
∗ Correspondingauthor.
E-mail:[email protected](J.Dou).
https://doi.org/10.1016/j.bjm.2017.04.014
Introduction
Polycyclic aromatic hydrocarbons (PAHs) are well known organic pollutants with 2–7 fused benzene rings and are speciallyrecognizedforpropertiesofcarcinogenicity, terato-genicityand mutagenicity.Typically,compoundswithmore aromaticringstructuresareassociatedwithhighertoxicity andposegreaterthreatstohumanbeings,animals,plantsand microorganisms.1 Amongthe 16priorityPAHslistedbythe
US Environmental Protection Agency (EPA), benzo(a)pyrene (BaP),withfivebenzenerings,wasnotableasthefirstreported carcinogen and is regarded as representative of PAHs.2
As a highly hydrophobic, non-volatile compound, BaP is degradation-resistantandmoreinclinedtoaccumulateinthe environmentovertime,whichleadstoconsiderable environ-mentalconcern.3PAHsarecommonhazardouscontaminants
inenvironmentalassessmentreportsandorganic pollutant-relatedstudiesandwidelyexistinsurfacewater,soilandeven groundwaterinmanyregionsofChina.Zhaoetal.4conducted
acomprehensiveinvestigationofPAHsinagriculturalsoilin thecoalproductionareasurroundingXinzhou,China.TheBaP equivalent ofsevencarcinogenic PAHs (majorcarcinogenic contributorsincludingBaP)accountedformorethan99%of the16PAHconcentrations.Liaoetal.5foundthattheBaP
con-centrationofcokingplantsoilscouldreach110±18.0mg/kg, whilethecontaminationdepthreachedmorethan10m.Wang etal.6notedapeakBaPconcentrationingroundwaterfroma
coalganguestackareaof46.4ng/L.Inaddition,becauseBaPis stronglyadsorbedontosoil,itexhibitsreducedmobilityand bioavailability,thusslowingbiodegradation,especiallyindeep soilwhereoxygenisscarce.RemediationmeasuresforBaP contaminationareurgentlyneededandtechnologiesforthe removalofBaPareinhighdemand.
Asan cost-effectivemethod, bioremediationtechniques do not require secondary treatment and can be applied atlargescales.7 BaP iseasilydegraded under aerobic
con-ditions in soil, and typical microorganisms reported to degrade BaP are white-rot fungi, Lasiodiplodia theobromae, algae,Rhodococcus, Alcaligenesdenitrificans, Sphingomonassp., and Stenotrophomonas maltophilia.8 However,oxygen is
gen-erallyinsufficientinreal casesduetoconsumption during thebiodegradationofaromaticswithfewerrings,leaving a considerableamountofBaPinanaerobicenvironments.9
Fur-thermore,anaerobicoranoxiclimitationscausegreatconcern forthedegradationofBaPindeepsoilandgroundwater.Thus, anaerobicbiodegradationofBaPisviewedasanimperative techniqueinpracticalsitecontaminationoperations.Recent studieshavedemonstratedthatBaPcanbedegradedwithout oxygenbymicroorganismsinriversedimentbyadding elec-tronacceptors,surfactantorco-metabolicsubstrates.10Thus
far,onlyaminorityofBaP-degradingbacteriaunder anaero-bicconditionshavebeenreportedtobeisolatedsuccessfully. Lianget al.11 isolatedonefacultativeanaerobePseudomonas
sp.JP1,whichcandegradeavarietyofPAHswith3–5rings except pyrene under nitrate-reducing conditions. The BaP removalratewasonly30%after40daysofcultivationata con-centrationof10mg/L.Intheresearchmentionedabove,the challengeofBaPanaerobicbiodegradationliesinthescarcity ofavailabledegradingbacteria,andtherefore,theisolationof
anaerobicdegradationbacteriahasbecomearesearchtopic ofgreatinterest.
Asacellulolyticandhemicellulolyticmicroorganism, Cellu-losimicrobiumsp.producesavarietyofdegradativeenzymes( -glucosidase,protease,glycosidehydrolaseandchitinase),and iswidelyappliedinbiodegradationofcellulosesandxylans andinalcoholicfermentation.12Naeemetal.13usedan
indige-nouschromium-resistantbacterialstrain A8 identifiedasa
Cellulosimicrobiumsp.intanneryeffluenttreatmentand98.6% ofCr(VI)wasreducedtothelesstoxicCr(III)formatthe con-centrationof900mg/L(witha1800mg/Ltolerancelimit), illus-tratingthatCellulosimicrobiumsp.hasastrongtolerancetohigh concentrationsofheavymetalions.Duetotheunique prop-ertiesoffibrolyticenzymes,Cellulosimicrobiumsp.couldbe uti-lizedinthebiodegradationofBaPunderanaerobicconditions. Microbialactivitiescanbeinfluencedinvariouswaysby the presenceofmetalions.Inparticular, heavymetalions mayhaveaperniciouseffectondegradingbacteriausedfor BaP biodegradation. Chenetal.14 investigatedthe
degrada-tionprocessofBaPinwateranddiscoveredthatdegradation abilitywasinhibitedwhentheconcentrationofcopper(Cu(II)) ionswaslower than 1mg/Lor higherthan10mg/L.Biswas etal.15studiedPAHbiodegradationbynativePAH-degrading
bacteria in a mixtureof heavymetals and concluded that bioavailablecadmium (Cd)was themainsourceoftoxicity for degrading bacteria. One metal-immobilizingorganoclay wasfoundtobeabletoreduce(Cd)toxicityandenhancePAH biodegradation.Additionally,temperature,time,solutionpH and availablecarbon sourceall affectthemicrobialactivity duringbiodegradation.Inthiswork,aBaP-degradingstrainof CellulosimicrobiumwasisolatedfromPAH-contaminatedsoilin acokingplantundernitrate-reducingconditions.The degra-dationperformanceundervariousenvironmentalconditions wasinvestigated.
Materials
and
methods
PreparationofPAHstocksolution
BaP,phenanthreneandpyrene werepurchasedfrom Sigma Chemicals, USA(purity>97%). ThePAHs were dissolved in acetone(CG,JTBakerUSA)andstockedataconcentrationof 1000mg/L.AllPAHstocksweresealedinambervials,wrapped withaluminumfoilandstoredat4◦Cpriortouse.
Preparationofculturemedium
Luria-Bertani(LB)mediumwasusedforbacterialenrichment and containedyeastextract (0.5%), tryptone(1%) and NaCl (1%).Mineralsaltmedium(MSM)wasusedasthedegradation solutionandcontained(g/L)Na2HPO4·10H2O(10.08),KH2PO4 (1.5), NH4Cl (1), NaNO3 (1), MgSO4·7H2O (0.2), FeSO4·7H2O (0.03), CaCl2 (0.02) and 1mL microelement solution. The composition of the microelement solution was as follows (mg/100mL):H3BO3(5.7),MnSO4·5H2O(4.3),ZnSO4·7H2O(4.3), CuSO4·5H2O(4.0),(NH4)6Mo7O24·4H2O(0.37),andCoCl2·6H2O (0.25).
IsolationandidentificationofBaP-degradingbacteria
Soil samples were collected at a depth of 40–80cm in a coking plant in Beijing that has been contaminated with PAHs for nearly 45 years. The total concentration of 16 USEPApriorityPAHsreached313.86mg/kg(dryweight,dw), includingphenanthrene(19.6%),anthracene(6.2%), fluoran-thene (26.3%), pyrene (18.1%), benzo(a)anthracene (6.5%), benzo(b)fluoranthene(8.5%),benzo(k)fluoranthene(3.0%),BaP (5.6%)andindeno(1,2,3-cd)pyrene(6.0%).BaPstockwasadded tothesoiltoguaranteeBaPasthedominantcontaminant, withafinalconcentrationof100mg/kg.Thesoilwassealed withpurenitrogen(99.9999%)followingacetoneevaporation andincubatedat25◦Cinthedark.BaP,asthesoleexternal car-bonsource,andnitrateastheelectronacceptorwereadded everysixmonthstoinduceBaPbiodegradationfor10years. ThesubsequentisolationtechniqueswereadoptedfromDou etal.16SoilsampleswithBaP-degradingbacteriaweremixed
withBaPsolution(25mg/L)andthebacteriaweretransferred intosupernatantfluidinthesoil-watermatrix.BaP-degrading strainsweretheninoculatedintoaBaP-supplemented MSM-agarplate(25mg/L)forfurtherpurification.
The isolated strains were identified using 16S rDNA sequenceanalysis.DNAwasextractedfrom isolated micro-bialcellsandamplifiedusingthepolymerasechainreaction (PCR).PCRamplificationwasconductedusingtheuniversal primers27F(5′-AGAGTTTGATCCTGGCTCA-3′)and1492R(5′ -TACCTTGTTACGACTT-3′) witha concentrationof 20M for both.17Eachamplificationreactionwascompletedusing25L
PCRMasterMix,2L27F,2L1492R,2LDNAlysate,and19L double-distilledH2O(ddH2O).Amplificationwasconducted for28cycles(94◦Cfor30s,55◦Cfor40s,and72◦Cfor90s)ina 100-seriesthermalcyclerafteraninitialdenaturationat94◦C for6min.ThePCRproductsweretestedbytheBeijingBiomed Co.,Ltd.The16SrDNAsequenceswerealignedwithpreviously publishedsequencesfromtheGenBankdatabaseofNational CenterforBiotechnologyInformation(NCBI)usingtheBLAST program(http://blast.ncbi.nlm.nih.gov).
Thegrowthcharacteristicsoftheisolatedstrain
BasedonitsBaPdegradationability,onestrainwasselected asthetargetstrainandusedforfurtherstudies.Thestrain was subject to biological and ecological characterization usingopticalmicroscopeandscanningelectronmicroscope observations.Duetotheimportanceofthecarbonsourcefor bacteriagrowth,variouscarbonsources(glucose,sucrose, sol-ublestarchandhigh-densitycornamylodextrins)wereadded totheMSMtoinvestigatethegrowthbehavioroftheisolated strain.Theresultswerecomparedwithits performanceon LBmedium.
Cultureconditions
Theisolatedstrainpreviouslycultivatedin100mLLBmedium at35◦Cfor24hwithashakingspeedof200r/minwas cen-trifugedat6000r/minfor15min.Thecellpelletswerewashed withMSM solution three timesto removeresidual LB and resuspended inMSM mediumand usedas inocula forthe followingBaP-degradationexperiments.
BaP-biodegradationexperiment
Biodegradationexperiments wereperformed indetest oxy-genbottles(100mL).ThePAHstocksolutioninacetonewas spikedintobottleswithMSM(20mL)broth,andthebottles were placedin a fume hood fortwo hours to allow com-pleteevaporationofacetone.Dissolvedoxygeninthebottle wasremovedbyaddingNa2S·9H2O(0.2g)afterthe evapora-tionofacetone.Theisolatedstrainwastheninoculatedinto themedium.Thebottleswerecappedwithsuba-sealsilicone septa,wrappedwithaluminumfoiltopreventphotolysisand incubatedat200r/minunderanaerobicconditions.Samples from individualbottlewerecollectedatthe desiredtimeto measureresidualBaP,andtheremovalratesofBaPwere cal-culatedtoelucidatetheoptimalconditions.Allexperiments wereconductedintriplicate,anderrorbarswerecalculatedas thestandarddeviationsofthemeasurements.
Theeffectsoftemperature,pH,BaPconcentrationand ini-tialbacterialmassandindividualmetalionsonBaPremoval ratesbytheisolatedstrainwereinvestigated.Theincubation temperatures wereset at20,25,30and 35◦C withainitial BaPconcentrationof25mg/L.AlsoataBaPconcentrationof 25mg/L,theinitialpHofthemediumwasadjustedto3,4,5, 6,7,8,9,10,11and 12byadding 0.1MNaOHor0.1MHCl. Five graded initialBaP concentrationsat 2.5,5.0, 10.0,25.0 and50.0mg/Lwerestudiedindividually.Theinitialbacterial masses wereregulated byadding variousamountsof inoc-ula.TheionconcentrationofFe(II),Zn(II),Cu(II),Mn(II),Hg(II), Co(II),Pb(II)andCd(II)was10mg/Lbyaddinginorganic com-pounds ofCo(NO3)2, FeSO4, MnCl2, CuCl2, PbCl2, Zn(NO3)2, HgCl2 andCdCl2with10mg/LBaPinthe medium,samples onlycontainBaPwereemployedascontrol.
Allexperimentsaboutconcentrationandmetalionswere conducted ina shakerat200r/min at35◦C with brothpH adjustedto7.Parallelsamplesindetestoxygenbottles with-outanyinoculationwereemployedasblankcontrol.
Theco-metabolic biodegradationbehaviorofBaP bythe isolatedstrainwasalsoinvestigated.Thereadilyutilizable car-bonsourceglucose,infiveserialconcentrations,andsuccinic acid,d-malic acid,l-malicacid,citricacidand tartaricacid
ataconcentrationof10mg/Lwereindividuallychosenas co-metabolicsubstrates.PAHswithsimilarstructuretoBaPwere addedtotestthecarbonsourceutilizationpatterns(priorities ofutilization)oftheisolatedstrain.Three-ringandfour-ring PAHs, phenanthrene and pyrene were used as representa-tivesfortheco-degradationstudies.Thetotalconcentrationof PAHswasadjustedto10mg/L(1.5mg/L,3.5mg/Land5mg/L forphenanthrene,pyreneandBaP,respectively).Theresults werecomparedwiththeBaPdegradationperformancewhen BaPwasthesolecarbonsource(BaPincontrol).
ResearchondegradationmechanismofBaP
The BaP metabolites during biodegradation process were extractedwithdichloromethaneafterbiodegradationof
ten-days. The extracts were dried over anhydrous sodium
sulfate, concentrated and filtered prior to analysis by gas chromatography–massspectrometry(GC–MS).18GC–MS
1.0mL/minwasusedasthecarriergas.Theoven tempera-tureprogramwasasfollows:100◦Cfor1min,thenlinearly increasedatarateof15◦C/minup to180◦C,180–310◦C at arampof5◦C/min,andmaintainedat310◦Cfor2min.The spiltflow was24mL/min,injector temperature was 250◦C. Themassspectrawererecordedat1scan/sunderelectronic impactwithelectronenergyof70eV.Massrangewasbetween m/z30–450.AlibrarysearchwascarriedoutbyusingNIST11.
AnalyticalmethodforPAHs
Samplesfrombottleswereextractedtwicewith20mL cyclo-hexaneeachtime(CG,JTBakerUSA).Ineachextraction,the mixturewas vigorouslyshakenfor10minandsubjected to ultrasonicextractionina25◦Cwaterbathat40Hzfor15min. Themixturewasallowedtophaseseparate,andtheorganic phasewascollected.Thetwoorganicsolventswerecombined, andresidualwaterwasremovedbyaddinganhydroussodium sulfate. The solution was further diluted to the appropri-ateBaPconcentrationwithHPLC-grademethanolandfiltered throughanylonMilliporemembranefilter(0.22m).ThePAHs wereidentifiedusingHPLCwithaUVdetectorat254nmanda C18reverse-phasecolumn(dimensions4.6mm×250mm,I.D., 5-mparticlesize);methanol(CG,JTBakerUSA)andultrapure water(90/10,v/v)wasusedwithaflowrateof1.0mL/minas themobilephase.Theinjectionvolumewas20L.Theentire processtook20min,andtheconcentrationofeachPAHwas calculatedfromthecurvebasedonthepeakareawithvarious knownconcentrations.
ThePAHdegradationrate(%)wascalculatedasfollows:
R= C0−Ct C0 ×100%
whereRrepresentstheremovalratios(%),andC0andCtare the initialandfinalPAH concentrations.All measurements werethemeanvaluesofthreereplicates.
Results
IsolationandcharacterizationofBaP-degradingstrain
A bacterial strain using BaP asits sole carbon and energy sourceundernitrate-reducingconditionswas isolatedwith plate-screeningtechniquesafterrepeateddomesticationand enrichment. The strain was identified using 16S rDNA sequenceanalysisandwasnamedCWS2.Duringthepartial 16S rDNAsequence determination,approximately656base pairsofthesequenceweredepositedintheGenBankdatabase.
Fig.1showsaphylogenetictreeandtheclosestrelativesof CWS2 listed according tothe GenBankdatabase. Basedon theresults,thestrainCWS2provedtohaveastrong relation-shipwithCellulosimicrobiumsp.TUT124(656bp,99%identical), Cellulosimicrobium sp.A24 (655bp,99% identical), and Cellu-losimicrobiumcellulans(655bp,99%identical).
The CWS2 colonieson solid plates were yellow, round, non-transparent,smooth,andwet witharegularedge,and thesurfaceofthecolonieswasuplifted.Asinglebacterium wasrod-shapedwithadiameterof0.2–0.3mandlengthsof 0.6–1.0m. Thestrainwas determined tobeGram-positive andcatalase-positive,anditsoxidaseactivitywasnegative. According to these characteristics and instructions from “Bergey’s Manual of Systematic Bacteriology,”19 the strain
CWS2 showed high similarity to Cellulomonas. Therefore, basedonthegeneticresults,thephysiologicaland
Cellulosimicrobium funkei strain 11 (KJ571073.1)
Cellulosimicrobium sp. TV30bNov(KJ482900.1)
Cellulosimicrobium cellulans(KJ513474.1)
Cellulomonas sp. MN 60.3(AJ313025.1)
C.cartae MSD 20106(EU816697.1)
Cellulosimicrobium sp. DICP-FYB-QH03-1(JN194192.1)
Cellulosimicrobium sp. 20.2 KSS(HE575928.1)
Cellulosimicrobium cellulans strain MWM-81 (JX535362.1)
Cellulosimicrobium sp. 20.1 KSS(HE575927.1)
Cellulosimicrobium sp. I20-1( EU181237.1)
Cellulosimicrobium funkei strain W6122(NR_042937.1)
Cellulosimicrobium sp. 139-5(GQ478418.1)
Bacterium WU13(KM264371.1)
Cellulomonas sp. DB-18(EU439413.1)
Cellulosimicrobium cellulans strain CA817(KF040973.1)
Cellulosimicrobium cellulans (AB098580.1)
Cellulosimicrobium sp. A24(KC434977.1)
Cellulosimicrobium sp. TUT1242(AB188222.1)
CWS2
71 55 47
47
23 20 47
42 32
0.0005
69 63 57 51 45 39 33 27 21 15 9 3 -0.2
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8
Absorbance (OD
600nm
)
time (h)
sucrose starch LB glucose corn amylodextrins
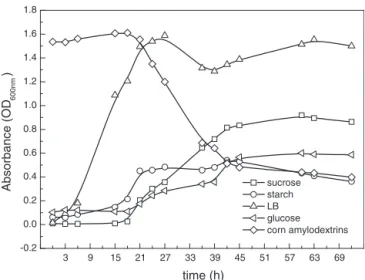
Fig.2–GrowthcurvesofCellulosimicrobiumcellulansCWS2
indifferentMSMmediums(sucrose,starch,glucoseand cornamylodextrins).Alltheexperimentalsettingsfollowed anaerobiccultivationrequirementsandthetemperature wascontrolledconstantlyat35◦C.Bacterialcellnumbers
areestimatedthroughtheopticaldensitymeasuredata wavelengthof600nm(OD600).Forthecurveofcorn amylodextrins,thehighertheOD600nmthehigher concentrationofcornamylodextrinsduetoitscolor,and theOD600nmdecreaseasthebacteriagrowusingthecorn amylodextrinsascarbonsource.
ical characteristics of the strain, the isolated strain CWS2 wasrecognizedasaCellulosimicrobiumsp.andwasnamedas C.cellulans CWS2.The genomic informationforC. cellulans CWS2 was deposited in the GenBank under the accession numberKX496338.
Tofurtherinvestigateitsgrowthbehavior,CWS2was culti-vatedonothercarbonsourcesincludingglucose,sucrose, sol-ublestarchandcornamylodextrins.Fromthegrowthcurves ofthestrainCWS2(Fig.2),differentlagphaseswereexhibited andthelongestwas16h.Specifically, CWS2begangrowing immediatelyfromthetimeofinoculationandthecell num-berreachedapeakwithin24hinLBmedium.A2ndgrowth periodwasobservedafterarelativelyshortattenuatedphase (between33hand42h).Whenstarchwasused,thecell num-berincreasedslowlyafter6handthecelldensitysignificantly increasedbetween15 and 24h. Amongall carbon sources, CWS2demonstratedthelongestlagphasewhencultivatedin glucose,solublestarchandcornamylodextrinmedia. Interest-ingly,theseallshowedasimilarlineargrowthperiodbetween 24and42hintheexponentialphase.Accordingtothe over-all growth behavior, LB and sucrose media provided more favorableenvironmentsforstrainCWS2,andthecelldensity indicatedhigherbacterialbiomass.ThecellbiomassofCWS2 culturedinLBmediumwasfourfoldhigherthanbiomass cul-turedinglucosemediumatthestationaryphase.Thus,LB mediumwasselectedforfurtherbacterialproliferation.
BiodegradationofBaP
A BaP concentrationof 25mg/L was adoptedin the inves-tigation oftemperature influenceon bacterialbehavior. As
showninFig.3A,nosignificantdifferencewasobservedinBaP degradationbehaviorat20and25◦Cwithin12days.BaPwas degradedfasteratboth30and35◦Cinthefirst5days com-paredtotheothertwotemperaturesettings,butdifferences indegradationspeedunderthetwoconditionswerewidening onthefifthday.ThedegradationrateofBaPat35◦Cwas18.0% higherthanthatat30◦Cafter12days.Thedegradationability at20or25◦Cwassignificantlylowerthan30and35◦Cduring thewholedegradationprocess.Thebiodegradationkineticsat 20and25◦Cfollowedazero-orderratemodel;theequations were y=6.96+3.19t, R2=0.9897(yisthe biodegradationrate (%)attimetandtisthedegradationtime)andy=10.60+2.56t, R2=0.977,respectively.Thedegradationcapacityat30◦Cand 35◦C obeyed first-order kinetics, and the equations were y=43.04–43.04e(−0.28t)(R2=0.9163)andy=62.10−62.10e(−0.20t) (R2=0.9290).Accordingtothefittingcurves,BaP biodegrada-tionratesusingCWS2below30◦Cremainedstableandwere notrelatedtotheBaPconcentrationremaininginthemedium. Thisindicatedthatdegradationabilitywasonlyconstrained bytemperature whenall other factors weresuitable under theexperimentalsettings.Onthefirstday,nearlyequivalent degradationrates(15%)wereobservedwhenthetemperature was25,30or35◦C.Thisindicatedthatthebacteriaexhibited similaradaptationabilityundertheseenvironments.Notably, almostthesamedegradationrate(between38.0%and42.8%) wasobtainedonthe12thdaywhenthetemperaturewas20, 25or30◦C.Whentemperatureexceeded30◦C,thehigher tem-peraturewascorrelatedwithhigherbiodegradationabilityfor CWS2,andthetotalamountofBaPdegradedalsoincreased. Basedontheexperimentalresults,CWS2culturedat35◦Cwas adaptedinsubsequentBaPdegradingtests.
AsshowninFig.3B,C.cellulansCWS2exhibitedhigherBaP biodegradation efficiencyunderneutraland mildlyalkaline conditions.HigherBaPdegradationrateswereobservedwhen thesolutionpHrangedfrom7to10,andtheBaP biodegrada-tionabilitybyCWS2proceededaspH10>pH9>pH8>pH7, indicatingthattoacertainextent,CWS2wasfavorably adapt-abletopHchanges.After10daysoftreatment,theBaPremoval ratewas58.7%atpH10,butonly50.0%atpH7.BaPremoval declinedsharplytobelow25.0%whenpHwaslowerthan6 duetorestrainedactivityofthedegradingenzyme.Incontrast, CWS2appearedtoadapttohighlyalkalineenvironmentsand obtainedabetterremovalratewithapHashighas12. Accord-ingtotheseresultsandthetypicalpHvaluesinnaturalwater environments,pH7wasselectedforthedegradationofCWS2 withhighefficiencyandlowcost.
Hg Cd Pb Zn Co Mn Fe Cu control 0 10 20 30 40 50 60 70 80
BaP degradation rates (%)
metal ions
(D)
12 11 10 9 8 7 6 5 4 3 2 1 0 10 20 30 40 50 60 70
20ºC 25ºC 30ºC 35ºC
BaP biodegration rates (%)
time (d)
(A) 3 4 5 6 7 8 9 10 11 12
0 10 20 30 40 50 60
BaP degradation rates (%)
pH
(B)
15 13 11 9 7 5 3 1 0 10 20 30 40 50 60 70 80 90
BaP degradation rates (%)
time (d) 2.5 mg/l 5.0 mg/l 10 mg/l
25 mg/l 50 mg/l
(C)
inocula
10 8
6 4
2 0
volume of inocula addition (ml)
Fig.3–EffectsofenvironmentalfactorsonBaPbiodegradationbyCellulosimicrobiumcellulansCWS2underanaerobic
conditions.(A)EffectoftemperatureonBaPbiodegradation.(B)EffectofsolutionpHonBaPbiodegradation.(C)Effectsof differentinitialconcentrationsanddifferentvolumeofinoculaadditiononBaPbiodegradation.(D)Effectofco-existing metalionsonBaPbiodegradation.TheinitialBaPconcentrationwas25mg/Lforallthebatchesin(A)and(B).The temperaturewasmaintainedat35◦Cforallexceptin(A),andsamplesweremeasuredatday10and13in(B)and(D),
respectively.ThesolutionpHwas7.0in(C)and(D).TheinitialBaPconcentrationwas10.0mg/Lin(D).Theerrorbars representthestandarddeviationsoftriplicatesamplemeasurements.
similarrisingtrendwithin13days;thesecurveswerenearly parallel. Thisis inaccord withthe typical bacterial degra-dation performance and indicated that the contaminants wereremovedatastableleveloncethemicroorganismswere fullyactivated.However,whencontaminantswerelimitedto below2.5mg/L,thebacteriadegradedthematahighinitial ratebut failed tomaintainthis rate duetothe limitations ofremaining BaP. The final BaP degradation rate obtained ataBaP concentration of2.5mg/Lwas 75.8%,close tothe highestrateof78.8%(initial BaPconcentration of10mg/L). The degradation curves at 50.0mg/L showed that CWS2 maintainedaconstantdegradationspeedevenatveryhigh BaPconcentrations;15mg/LofBaPwasconsumedin15days. Itispredictedthatthepacecanbecontinuouslymaintained asexhibitedinthelow-BaPconcentrationcurves.Thus,strain CWS2possessedastrongBaP-degradationability.
Degradation tests were carried out ata BaP concentra-tionof10.0mg/Lat35◦CandpH7for15days.Thebiomass dosages added at 20mL MSM were 0.25,1.25, 2.5, 5.0 and 10.0mL.TheresultsinFig.3Cindicatethatbiomasscouldbe amajorlimitingfactorinBaPbiodegradationandshouldbe properly studiedwhenbacteriaare usedtoremediate BaP-contaminatedsoilorwater.
concentrationsofthoseionsresultinserioussideeffects.As showninFig.3D,BaPbiodegradationwasinhibitedbyCu(II), Mn(II)andCo(II)inalimitedrange(removalratiosdecreased below10%).WhilethestrainCWS2demonstratedtolerance toFe(II)andZn(II).Thestrainexhibitedobviouslyhigher tol-erance to Zn(II) due to the fact that the BaP removalrate increasedundercurrentexperimentalsettings.Inthisstudy, noobviousinhibitioneffectonBaPdegradationwasobserved withtheadditionofPb(II)andHg(II).
EffectofadditionalcarbonsourcesonBaPbiodegradation
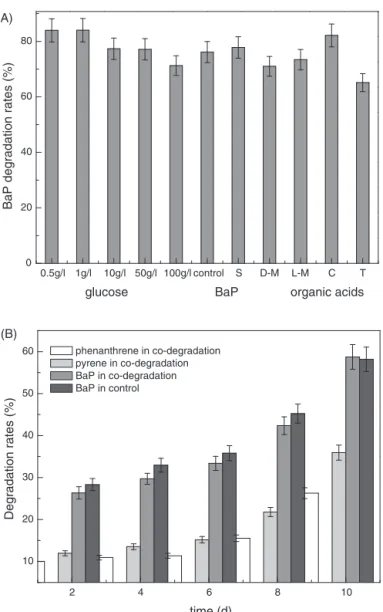
Asacommonenergeticmaterial,glucoseiseasily biodegrad-ableandisutilizedpreferentiallybybacteriainthepresence ofvariouscarbonsources,particularlyinanaerobic environ-mentssuchasfermentationconditions.Accordingly,glucose isfrequentlyusedasaco-substratetopromotemicroorganism growthfordegradationoforganicpollutantsandhasalsobeen widelyusedasaco-substrateforanaerobicbiodegradationof PAHs.10Asdiscussedabove,strainCWS2begantothriveafter
alagphaseof16hinaglucosesubstrate,andthebacteria reachedmaximumbiomassin45h.AsillustratedinFig.4A, theproperadditionofglucosecanenhanceBaPdegradation. Glucose atconcentrationsof 0.5and 1.0g/L had apositive effectonBaP biodegradation,and theremovalefficiencyof BaPincreasedby10% comparedwiththe control.However, noevidentincreaseinBaP removalefficiencywasobserved whentheglucoseconcentrationwasabove10g/L.BaP degra-dationwasslightlyinhibitedoncetheglucoseconcentration surpassed50g/L.Thus,whenglucose servedasthecarbon sourceforbacterialgrowth,properadditioncanenhanceBaP biodegradation.LMWorganicacidsareidealcarbonsources formostmicroorganisms, astheyare metabolic intermedi-atesofvariouscarbonsources.ItisobservedinFig.4Athat theaddition ofLMWorganic acidshad differenteffectson thedegradation ofBaP.Relatively, citricacidpromoted BaP biodegradation,whiletartaricacidslightlyinhibited degrada-tion.Otherorganicacids(succinicacid,d-malicacid,l-malic
acidandtartaricacid)influencedBaPdegradationinalimited range.
InFig.4B,phenanthreneandpyreneweredegradedto dif-ferentdegrees byCWS2.However,the degradation ratesof phenanthreneweresignificantlylowerthanthatforpyrene, andbothweremuchlowerthan theBaPdegradationrates. Incomparison,theBaPdegradationcapabilityofstrainCWS2 revealednosignificantdifferenceinmixedPAHssolutionor BaP-onlysolution.Thus,weconcludedthatstrainCWS2had ahighselectivityforBaPamongmixedPAHs.The biodegrada-tionratesofphenanthreneandpyreneremainedunchanged inthefirst6daysandthenincreased.Thedegradation pat-terns of BaP under both conditions were similar, as the degradationrateincreasedfromapproximately26%atthe sec-onddaytoapproximately59%atthetenthday.
DegradationmechanismofBaP
The qualitative analysis of the metabolic
intermedi-ates of BaP biodegradation showed that some new
peaks were turned up in the chromatograms of samples compared with that of blank control. Several
identi-cal products were detected based on the GC–MS library
search and they were pyrene, 1-aminopyrene,
phenan-threne, 1-methylphenanthrene, 1,7-dimethylnaphthalene, 1-(2-hydroxypropyl)naphthalene, 1-methylnaphthalene, 2-hydroxy-3-(3-methyl-2-butenyl)-1,4-naphthalenedione, diethyl phthalate, and 2-acetyl-3-methoxybenzoc acid. A possiblebiodegradationpathwayofBaPwasproposedbased on the catalysisofenzymes todamagerings structuresas
Fig.5shown.
In our test, pyrene and 1-aminopyrene were the initial intermediate products detectedwhich were not generated from thefirstopeningofBaP,andtherelationshipbetween pyreneand1-aminopyrenewerestillunclear.Phenanthrene and1-methylphenanthrenewereintermediateproductswith three rings. Though four products with two rings were detected,itwasdifficulttofigureouthowtheyweredegraded. Finally,thesecompoundswerebrokendowntosomebenzene compoundswithmethoxyandcarboxyl.
Discussion
IsolationandcharacterizationofBaP-degradingstrain
The isolated strainwas initially identifiedas Cellulosimicro-biumsp.,andsomerelatedstudiesondegradationoforganics havebeen reported.Liu etal.20 isolatedastrain capableof
high phenol tolerance from rhizosphere soil in a sewage-treatment plant, and idealdegradation rates were reached when the phenol concentrationwas 500–1500mg/L. Analy-sis basedon 16S rDNA gene sequences demonstrated that thestrainbelongedtoC.cellulans.DavolosandPietrangeli21
studiedbacterialstrainsinsamplesfromaPAHswastewater treatmentinItalyanddetectedCellulosimicrobium.The Cellu-losimicrobiumspecieslistedaboveallshowedsimilarbehaviors asthestrainweidentified,thusthestrainCellulosimicrobium hasthepotentialtodegradePAHs.
BiodegradationofBaP
Temperature and pH affect not only the physiological growth of microorganisms but also the activity of their enzymes.Althoughmicroorganismscansurviveunderwide
temperature and pH ranges (even in extreme
environ-ments),appropriatetemperatureandpHconditionsmustbe determined to maximize growth and related physiological activities. In this way, functional microbes can be thor-oughlyutilizedinindustrialapplications.Asobservedinthe BaP degradation by strain CWS2, the pH of the medium decreased duetomicrobial productionofacid metabolites. Thesubstantialaccumulationoflow-molecular-weight(LMW) organicacidsfromsourcesofmicrobialmetabolismchanged pH markedly and influenced metabolic pathways.22 These
metabolicpathwayswerealsoaffectedbycarbonsources,and accumulationofmetaboliteswasalsorelatedtotheamountof carbonsources.Thus,thetypeandconcentrationofsubstrate playedanimportantroleinthegrowthofmicroorganisms.
T C L-M D-M S control 100g/l 50g/l 10g/l 1g/l 0.5g/l 0 20 40 60 80
BaP degradation rates (%)
glucose BaP organic acids (A)
10 8
6 4
2 10 20 30 40 50 60
Degradation rates (%)
time (d)
phenanthrene in co-degradation pyrene in co-degradation BaP in co-degradation BaP in control
(B)
Fig.4–Effectsofco-substratesonBaPanaerobicbiodegradationbyCellulosimicrobiumcellulansCWS2.TheinitialBaP
concentrationwas10mg/L.Theconcentrationsofphenanthreneandpyreneaddedintothemediumwere2.5and7.5mg/L, respectively.TheS,D-M,L-M,CandTrepresentedsuccinicacid,d-malicacid,l-malicacid,citricacidandtartaricacid,
respectively.Theerrorbarsrepresentthestandarddeviationsoftriplicatesamplemeasurements.
theycanbeharmfulorevenlethaltomicroorganismswhen impropermetalionsbindtotheenzyme,resultingin inac-tivation and denaturation. The biodegradation of BaP by microorganismsinasolutionistypicallycompletedbytwo steps:absorption tothe microbial cells surface, and trans-portintocellsfollowedbymetabolism.23Theuptakeofheavy
metals by organisms operates via the same mechanism: adsorptionofmetalionsonthecellsurfaceand intracellu-larbindingofheavymetalstothiol-containingcompounds. Theresultsfrom Chenet al.24 indicated that theactivities
ofmanganeseperoxidaseandligninperoxidasewere gener-allyinhibited byCd. Cd(II)iscytotoxic tocells anddirectly and severelyimpacts enzymatic activities involved in bac-terial metabolism. Chen et al.25 investigated the removal
mechanisms ofBaP withCu(II) atthe cellular interface of Stenotrophomonasmaltophiliaandconcludedthatthecellwall wasthemainadsorptionsiteforCuandfunctionedasa bar-rierforthetransportationofBaP.Astudyofmechanismsof
Cu- and Zn-binding using Pseudomonas putidaCZ1 through chemicalmodificationofbiomassbyChenetal.26indicated
that carboxylfunctional groupsin thecell wallsplayedan importantroleinthebindingofmetal.Thus,thereisahigh probabilitythatacidicmetabolitesofthemicrobeswith car-boxyl groupsassisted inCu(II) adsorptionand reduced the transportofBaP.InthemetabolismofBaP,theremustbe inter-mediatescontainingcarboxylgroupsfollowed bydecreased BaPdegradationrates.WhiletheheavymetalsHg(II)andPb(II) will denatureproteins and leadto a lossofvitality of the microorganisms.27
EffectofadditionalcarbonsourcesonBaPbiodegradation
microorgan-Benzo(a)pyrene
NH2
1-aminopyrene pyrene
Phenanthrene
OH
1-(2-hydroxypropyl)-naphthalene,
1,7-dimethyl-naphthalene 1-methyl-naphthalene,
O O
OH
2-hydroxy-3-(3-methyl-2-butenyl)
-1,4-naphthalenedione
O
OH O
O
acid benzoic 2-acetyl-3-methoxy
O O O
O
phthalate diethyl
1-methylPhenanthrene
Fig.5–ProposedpathwaysforanaerobicdegradationofBaPbyCellulosimicrobiumcellulansCWS2.
ismsmainly through two routes: consumption as the sole carbonandenergysourceorasanalternativecarbonsource. RecentstudiesshowedthatBaPcouldbedegradedbymixed bacteria, but most indicatedthat BaP isdegraded as a co-metabolicsubstrateunderanaerobicconditions.9Ambrosoli
etal.10reportedthatphenanthreneandpyrenedegradation
abilitywere enhancedremarkablywhen1g/kgglucosewas added asa co-substrate.Ye et al.29 drew anopposite
con-clusion:sucroseexertedasuppressiveeffectonanthracene biodegradation.Asstatedabove,strainCWS2effectively util-izesBaPasasole carbonandenergysource.However,both wereobservedinourexperimentalsettings,whichillustrated that the amount of glucose was the primary determinant onwhetherBaPbiodegradationwillbeenhancedbyglucose addition.
Thebiodegradabilityoforganicsismainlydependenton thecomplexityoftheirchemicalstructuresandcorresponding physicochemical properties. Organics with simpler struc-tureswillbeeasilyused.Organic acidsinour experiments hadsimilar structures.Thecomplexityorder ofthecarbon
sources is citric acid<tartaric acid<malic acid<succinic acid. However, the biodegradation ratios for strain CWS2 obtained from these acids did not follow this order. As discussedabove,BaPbiodegradationratesbyCWS2 dramat-ically decreased when the pH of the medium was below 10.Theorder ofpHvaluesforthesesolutionswas tartaric acid<citricacid<malicacid<succinicacid,whichwas pos-itivelycorrelatedwithdegradationrates(exceptcitricacid). Thus, citric acid addition contributed to promote the BaP degradation.
ThemolecularstructureofPAHsdeterminestheir stabil-ity and hydrophobicity and is the maininternal factor in controllingbiodegradability.Generally,LMWPAHstendtobe moreeasilybiodegradedthanhigh-molecular-weightPAHs.30
bothPAHsweredetectedinlimitedamountsinstudiesofBaP biodegradationbehaviorbyCWS2.
DegradationmechanismofBaP
Previous study have reported that naphthalene, phenan-threne,pyreneandother PAHswere completelyoxidizedto carbon dioxidebymixed bacteriathrough denitrification.31
While researches on degradation of PAHs by single
strain bacteria under anaerobic conditions were sel-domly reported, particularly the biodegradation of HMW PAHs. Liang et al.11 firstly investigated the degradation
pathway of BaP by Pseudomonas sp. JP1 under
anaero-bic conditions. He pointed out that the first opening of BaP ring generated 1,12-dimethyl-benzo(a)anthracene, 7,8,9,10-tetrahydrobenzo(a)pyrene, and 5-ethylchrysene, and then further degraded to pyrene, phenanthrene and 1-methylphenanthrene.Qinetal.32alsoproposedthe
anaer-obic degradation pathway of BaP by Microbacterium sp., the products were 4,5-dihydrobenzo(a)pyrene and 7,8,9,10-tetrahydrobenzo(a)pyreneduringtheprocessoffirstopening of BaP ring. In our test, only pyrene, phenanthrene and 1-methylphenanthrenewerethe sameintermediates found in the above pathway, thus the degradation pathway was differentfromthePseudomonassp.JP1.
Conclusion
C.cellulansCWS2wasisolatedfromPAH-contaminatedsoilin acokingplant.ThestrainusesBaP asthesole carbonand energysourceunder nitrate-reducingconditions. Tempera-tureandpHofthebrothwerethemainfactorsaffectingBaP removal,followedbytheBaPconcentration.ThehighestBaP removalrate(78.8%)wasobservedin13dayswhentheinitial BaPconcentrationwas10mg/L,andthestraindegradedBaPat aconstantrateevenatahigherconcentration(50mg/L).Inthe metalexposureexperiment,onlyCd(II)significantlyinhibited biodegradationamongothermetalions.Appropriateaddition ofglucoseenhancesBaPbiodegradation,whilestrongerLMW organicacids reducedegradationability.Phenanthreneand pyrenewere alsodegradedtovariousextentsbythestrain inthepresenceofBaP.Lastly,apossibledegradation path-wayofBaPunderanaerobicconditionswasproposed.Thus, thestrainexhibitedastrongBaPdegradationcapabilityunder naturalconditionsandhasgreatpotentialintheremediation ofBaP-/PAH-contaminatedenvironments.
Conflicts
of
interest
Theauthorsdeclarethattheyhavenoconflictofinterest.
Acknowledgements
This work was supported by the National Natural Sci-enceFoundation ofChina(51579010)andthe Fundamental ResearchFundsfortheCentralUniversities(2014NT32).
r
e
f
e
r
e
n
c
e
s
1.DamienB,SimonL,VincentB,etal.Occupationalexposure topolycyclicaromatichydrocarbons:relationsbetween atmosphericmixtures,urinarymetabolitesandsampling times.IntArchOccupEnvironHealth.2015;88:1119–1129.
2.IARC.Somenon-heterocyclicpolycyclicaromatic
hydrocarbonsandsomerelatedexposures.IARCMonographs
onEvaluationofCarcinogenicRiskstoHuman.vol.92.Lyon,
France:InternationalAgencyforResearchonCancer;2010.
3.RafinC,VeignieE,FayeulleA,SurpateanuG.Benzo[a]pyrene degradationusingsimultaneouslycombinedchemical oxidation,biotreatmentwithFusariumsolaniand cyclodextrins.BioresourTechnol.2009;100:3157–3160.
4.ZhaoL,HouH,ShangguanYX,etal.Occurrence,sources, andpotentialhumanhealthrisksofpolycyclicaromatic hydrocarbonsinagriculturalsoilsofthecoalproductionarea surroundingXinzhou,China.EcotoxicolEnvironSaf.
2014;108:120–128.
5.LiaoXY,MaD,YanXL,YangLS.Distributionpatternof polycyclicaromatichydrocarbonsinparticle-sizefractions ofcokingplantsoilsfromdifferentdepth.EnvironGeochem
Health.2013;35:271–282.
6.WangXW,ZhongNN,HuDM,LiuZZ,ZhangZH.Polycyclic aromatichydrocarbon(PAHs)pollutantsingroundwater fromcoalganguestackarea:characteristicsandorigin.
WaterSciTechnol.2009;59(5):1043–1051.
7.BishtS,PandeyP,BhargavaB,SharmaS,KumarV,Sharma KD.Bioremediationofpolyaromatichydrocarbons(PAHs) usingrhizospheretechnology.BrazJMicrobiol.
2015;46(1):7–21.
8.Machín-RamírezC,MoralesD,Martínez-MoralesF,OkohAI, Trejo-HernándezMR.Benzo[a]pyreneremovalby axenic-andco-culturesofsomebacterialandfungalstrains.Int
BiodeteriorBiodegrad.2010;64:538–544.
9.MeckenstockRU,MichaelS,ChristianG.Anaerobic degradationofpolycyclicaromatichydrocarbons.FEMS
MicrobiolEcol.2004;49:27–36.
10.AmbrosoliR,PetruzzelliL,MinatiJL,MarsanFA.Anaerobic PAHdegradationinsoilbyamixedbacterialconsortium underdenitrifyingconditions.Chemosphere.
2005;60:1231–1236.
11.LiangL,SongXH,KongJ,ShenCH,HuangTW,HuZ. Anaerobicbiodegradationofhigh-molecular-weight polycyclicaromatichydrocarbonsbyafacultativeanaerobe
Pseudomonassp.JP1.Biodegradation.2014;25:825–833.
12.YuanY,HuYB,HuCX,etal.Overexpressionand characterizationofaglycosidehydrolasefamily1enzyme
fromCellulosimicrobiumcellulanssp.21anditsapplicationfor
minorginsenosidesproduction.JMolCatalBEnzym. 2015;120:60–67.
13.NaeemA,BatoolR,JamilN.Cr(VI)reductionby
Cellulosimicrobiumsp.isolatedfromtanneryeffluent.TurkJ
Biol.2013;37:315–322.
14.ChenSN,YinH,YeJS,PengH,ZhangN,HeBY.Effectof copper(II)onbiodegradationofbenzo[a]pyreneby
Stenotrophomonasmaltophilia.Chemosphere.2013;90:1811–1820.
15.BiswasB,SarkarB,MandalA,NaiduR.Heavy metal-immobilizingorganoclayfacilitatespolycyclic aromatichydrocarbonbiodegradationin
mixed-contaminatedsoil.JHazardMater.2015;298:129–137.
16.DouJF,LiuX,DingAZ.Anaerobicdegradationof
naphthalenebythemixedbacteriaundernitratereducing conditions.JHazardMater.2009;165:325–331.
domesticwastewatersludgeforlandspread.WaterRes. 2012;46(8):2488–2504.
18.BinetP,PortalJM,LeyvalC.ApplicationofGC–MStothe studyofanthracenedisappearanceintherhizosphereof ryegrass.OrgGeochem.2001;32:217–222.
19.JohnGH,KriegNR.Bergey’sManualofDeterminative
Bacteriology.9thed.Baltimore:Williams&Wilkins;1994.
20.LiuAH,WangMM,YuanG,etal.Isolation,identificationand degradationcharacteristicsofaphenol-degrading
Cellulosimicrobiumsp.JSafEnviron.2013;13:24–28.
21.DavolosD,PietrangeliB.Molecularandphylogenetic analysisonbacterialstrainsisolatedfromaPAHs wastewatertreatmentplant.CurrResTopApplMicrobiol
MicrobBiotechnol.2009:313–316.
22.SilvaIS,GrossmanM,DurrantaLR.Degradationofpolycyclic aromatichydrocarbons(2–7rings)undermicroaerobicand very-low-oxygenconditionsbysoilfungi.IntBiodeterior
Biodegrad.2009;63:224–229.
23.KallimanisA,FrillingosS,DrainasC,KoukkouAI.Taxonomic identification,phenanthreneuptakeactivity,andmembrane lipidalterationsofthePAHdegradingArthrobactersp.strain Sphe3.ApplMicrobiolBiotechnol.2007;76:709–717.
24.ChenAW,ShangC,ZengGM,etal.Extracellularsecretionsof
PhanerochaetechrysosporiumonCdtoxicity.IntBiodeterior
Biodegrad.2015;105:73–79.
25.ChenN,YinH,TangSY,PengH,LiuZH,DangZ.Metabolic biotransformationofcopper-benzo[a]pyrenecombined
pollutantonthecellularinterfaceofStenotrophomonas
maltophilia.BioresourTechnol.2016;204:26–31.
26.ChenXC,ShiJY,ChenYX,etal.Determinationofcopper bindinginPseudomonasputidaCZ1bychemicalmodifications andX-rayabsorptionspectroscopy.ApplMicrobiolBiotechnol. 2007;74:881–889.
27.PetkovˇsekSAS,KopuˇsarN,TomeD,KryˇstufekB.Risk assessmentofmetalsandPAHsforreceptororganismsin differentlypollutedareasinSlovenia.SciTotalEnviron. 2015;532:404–414.
28.LilyMK,BahugunaA,DangwalK,GargV.Degradationof benzo[a]pyrenebyanovelstrainBacillussubtilisBMT4i (MTCC9447).BrazJMicrobiol.2009;40:884–892.
29.YeJS,YinH,QiangJ,etal.Biodegradationofanthraceneby
Aspergillusfumigatus.JHazardMater.2011;185:174–181.
30.WangLC,LinLF,LaiSO.Emissionsofpolycyclicaromatic hydrocarbonsfromfluidizedandfixedbedincinerators disposingpetrochemicalindustrialbiologicalsludge.J
HazardMater.2009;168:438–444.
31.NiemanJKC,SimsRC,McleanJE,SorensenD.Fateofpyrene incontaminatedsoilamendedwithalternateelectron acceptors.Chemosphere.2001;44:1265–1271.
32.QinW,ZhuY,FanFQ,etal.Biodegradationofbenzo(a)pyrene
byMicrobacteriumsp.strainunderdenitrification: