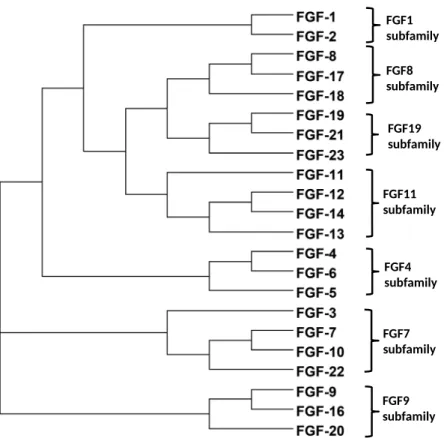
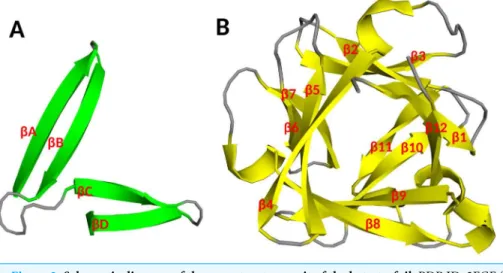
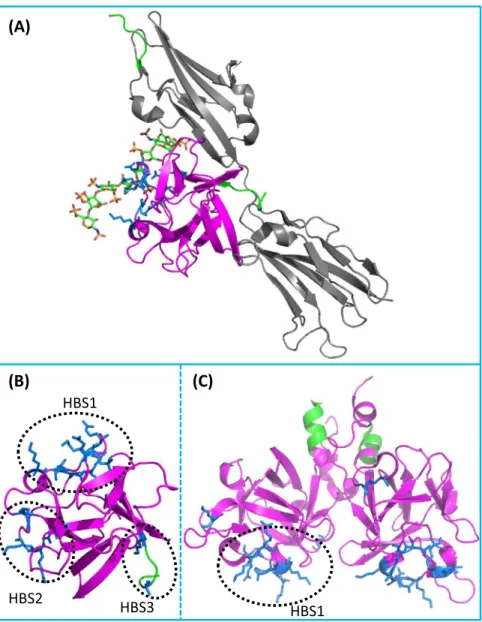
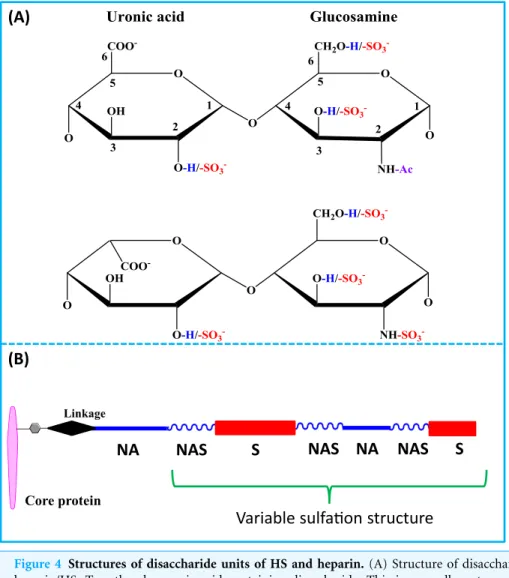
Fibroblast growth factors as tissue repair and regeneration therapeutics
Texto
Imagem
Documentos relacionados
The inner diameter of the iliac artery was significantly and similarly reduced in the SHR, SHR + molsidomine, and SHR + PETN groups compared to control Wistar rats.. The calculated
For the regeneration process related to the body fragment, group-2 presented better tissue architecture (3 minutes of laser exposure) compared to group-1 and the
MDA levels in testis tissues was significantly increased in the lead nitrate and sodium selenite plus lead nitrate treated groups compared to the control group,
A significant increasing in CAT activity was observed in the lead nitrate and mercury chloride treated groups compared with the control group.. Also significant
Compared with the untreated animals, the CD68+ macrophage population was significantly decreased over time in the wounds of treated animals, except on D20, when
Throughout the course of the regeneration process there was reduction of the CTDA in the control group (C21 and C21L) and in group D21L when compared to the respective 7-day groups
Wound inflammation-related miRNAs; digit wound healing and regeneration; regenerative myogenesis; rejuvenating muscle stem cells; immune control; post-natal and adult
sativum-treated group exhibited a significantly higher mast-cell and vascular distribution compared to animals in the Cicalfate, placebo, and control-sham groups. Thus, we
