Generation mean analysis for quantitative traits in sesame
(
Sesamum indicum
L.) crosses
Vijayarajan Sharmila, Siruthaiyur Krishnamoorthy Ganesh and Mahalingam Gunasekaran
Department of Plant Breeding and Genetics, Agricultural College and Research Institute, Madurai,
Tamil Nadu, India
Abstract
To study the nature and magnitude of gene effects for yield and its components in sesame (Sesamum indicum L.) we carried out generation mean analysis using the following four crosses of different sesame cultivars: VS 9510 x Co1; NIC 7907 x TMV 3; Cianno 13/10x VRI 1; and Si 1115/1 x TMV 3. The P1, P2, F1, F2, BC1and BC2of these generations were studied for seven quantitative traits. The analysis showed the presence of additive, dominance and epistatic gene interactions. The additive dominance model was adequate for plant height in the NIC 7907 x TMV3 and Si 1115/1x TMV 3 crosses and for capsule length in the VS 9510 x Co1, NIC 7907 x TMV 3 and Si 1115/1 x TMV 3 crosses. An epistatic digenic model was assumed for the remaining crosses. Duplicate-type epistasis played a greater role than complementary epistasis. The study revealed the importance of both additive and non-additive types of gene action for all the traits studied.
Key words:Sesame, quantitative trait, scaling tests, gene effects, bi-parental mating.
Received: December 13, 2005; Accepted: July 28, 2006.
Introduction
Sesame (Sesamum indicumL.) is an ancient and im-portant oil seed crop also known in India as til, ellu, beniseed, simsim and Sesame is often referred to by the epi-thet “the queen of oil seeds ” because it is prized not only for its nutritive value but also for the quality and quantity (44% to 52%) of its oil which is rich in vitamin E and has a significant amount of linoleic acid that can control blood cholesterol levels. The protein content of sesame seed is about 26.25% and the sulfur-containing amino acid methionine is present at a concentration of about 3.4% (Mosjidis, 1982). Sesame seed oil is resistant to oxidation and rancidity under normal storage conditions. Production of sesame hybrids can be accomplished either genetically using the cytoplasmic male sterility system, which have not been yet reported in sesame, or non-genetically by manual emasculation and crossing for the production of hybrids, which is the preferred route due to the fact that sesame has epipetalous stamens. Emasculation is the simplest crossing technique for producing sesame F1 hybrids it might be fea-sible to use this technique to produce hybrids and obtain a quantum jump in yield with minimal cost. In practice,
how-ever, this technique has not contributed much to the current oil seed scenario, with the average sesame productivity in India being practically stagnant during last few years and not impressive as compared to other sesame growing coun-tries of the world. So the challenge of the Indian sesame breeder is to step up yield through genetic improvement by adopting different breeding methodologies. For genetic im-provement of the crop, the breeding method to be adopted depends mainly on the nature of gene action involved in the expression of quantitative trait. Line x Tester (L x T) analy-sis is used to select the parents based on their combining ability but fails to detect the epistasis, which remains the most complex problem and on which it is extremely diffi-cult to obtain reliable results. The presence or absence of epistasis can be detected by the analysis of generation means using the scaling test, which measures epistasis ac-curately whether it is complimentary (Additive x Additive) or duplicate (Additive x Dominance) and (Dominance x Dominance) at the digenic level. The objectives of this re-search described in this paper were to obtain information on the nature of gene action in sesame to provide a basis for an evaluation of selection methods for the improvement of the sesame population.
Material and Methods
We used eight Sesamum indicum L. cultivars (VS 9510, Co 1, NIC 7907, TMV 3, Cianno 13/10, VRI 1, Si
Send correspondence to V. Sharmila. Senior Research Fellow, De-partment of Pulses, Centre for Plant Breeding and Genetics, Tamil Nadu Agricultural University, Coimbatore, 641 003 Tamil Nadu, In-dia. Email: [email protected].
1115/1 and TMV 3) provided by Regional Research Sta-tion, Vridhachalam, Tamil Nadu, India. The experiments involved the six basic generations (the P1 and P2 parent cultivars, the F1and F2first and second filial generations, and the BC1and BC2first and second back crosses) of four combinations of the parental cultivars, these combinations being VS 9510 x Co 1, NIC 7907 x TMV 3, Cianno 13/10 x VRI 1 and Si 1115/1 x TMV 3. The segregating and non-segregating parental populations were cultivated in a Randomized block design with two replications at the Agri-cultural College and Research Institute, Madurai, India during the winter of 2003. We used the parents of the re-spective crosses as the male parent and the F1generation as the female parent and effected back crosses to produce the B1(F1back crossed to P1) and B2(F1back crossed to P2) generations and the F1 hybrids were selfed to obtain F2 seeds. All these generations were produced during two cropping seasons and, as such, all the six generations had to be grown together during the same cropping season. The row-length was always three meters but the number of rows varied as follows: three rows, for the non-segregating P1, P2 and F1; 40 rows for the F2; and 20 rows for the BC1and BC2 generations. Since the non-segregating generations repre-sent the homogeneous population while the segregating generations represent the heterogeneous population the sample size (i.e.number of plants analyzed) varied as fol-lows: 30 plants for the P1, P2and F1generations; 400 plants for the F2generations; and 200 plants in the BC1and BC2 generations. The traits assessed were days to maturity, plant height (cm), number of branches per plant, number of capsules per plant, capsule length (cm), weight (g) of 1000 seeds and seed yield per plant (g).
The mean values, standard errors and variances of the different generations were subjected to weighed least-squares analysis using the scaling test (Mather 1949) and the joint scaling test to estimate gene effects. The ge-netic effects were estimated using the models suggested by Mather and Jinks (1971) and Jinks and Jones (1958). The significance of the scales and gene effects were tested by using the t-test (Singh and Chaudhary, 1985).
The A, B and C scaling tests were carried out for seven traits indicated the presence of non-allelic interac-tions in almost all cases. The A and B scaling tests provided the evidence for the presence of additive x additive (i), ad-ditive x dominance (j) and dominance x dominance (l) type gene interactions. The C scaling test provided a test for type l epistasis. The type of epistasis was determined only when dominance (h) and dominance x dominance (l) effects were significant, when these effects had the same sign the effects were complementary while different signs indicated dupli-cate epistasis (Kearsey and Pooni, 1996).
Results and Discussion
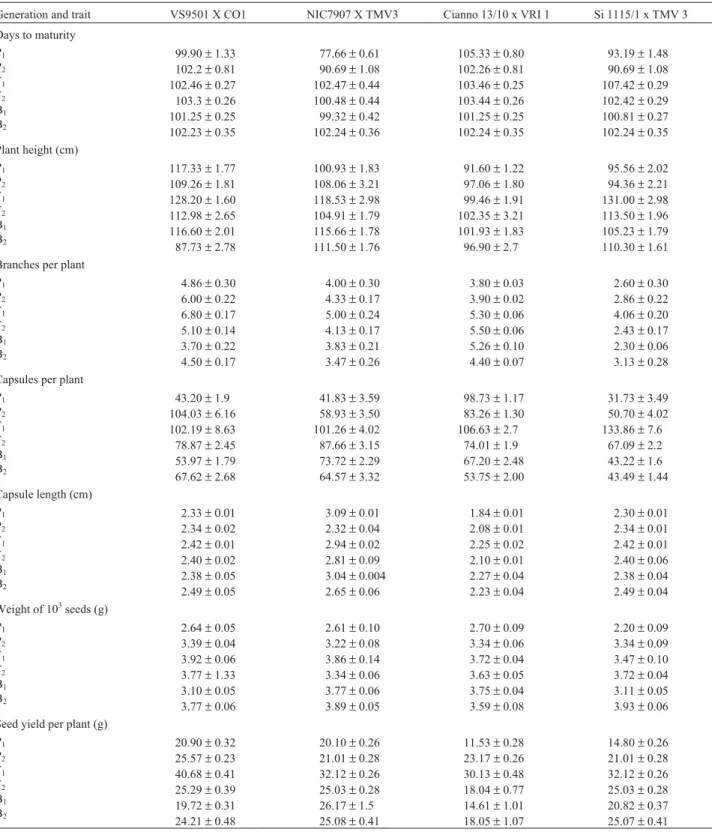
The mean and standard error of the six generations with four crosses for seven traits are presented in Table 1,
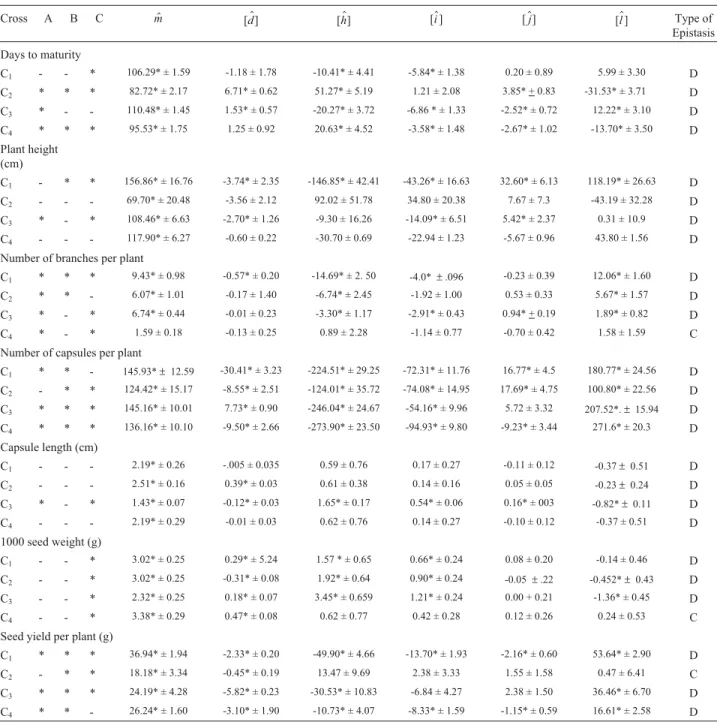
with the means values for the scaling joint scaling tests and their interaction effects being presented in Table 2.
The hybrids performed better than their respective parents in all crosses except for the VS 9510 x Co 1 cross in regard to the number of seed capsules per plant and for the NIC 7907 x TMV 3 cross in regard to capsule length, both these crosses showing inferior performance than their re-spective P1generations. In general, however, the trait mean values for the F1and F2generations were higher than the corresponding values for the BC1 and BC2 generations, while the values for the F2generation were lower than the corresponding values for the F1generation.The mean per-formance of the BC2segregating generation was lower than that of the BC1generation for all crosses and traits except for plant height and number of branches per plant for the Si 1115/1 x TMV 3 cross, number of capsules per plant and capsule length for the VS 9510 x Co 1cross, and seed yield per plant and the weight in grams of 1000 seeds for the VS 9510 x Co 1, Cianno 13/10 x VRI 1 and Si 1115/1 x TMV 3 crosses. The expected mean (m) of the four possible homo-zygotes was positive and significant in all the crosses for all the traits.
A simple additive/dominance model was adequate as inferred from the non-significance of all the scales for plant height in the NIC 7907 x TMV 3 and Si 1115/1 x TMV 3 crosses, capsule length for the VS 9510 x Co 1, NIC 7907 x TMV 3 and Si 1115/1 x TMV 3 crosses, and seed yield per plant for the NIC 7907 x TMV 3 cross. For the remaining crosses, an epistatic digenic interaction was found to be a suitable fit, since the scaling and/or joint scaling tests were significant.
The additive, dominance and epistatic types of gene interaction in each cross for different trait were found to be different from each other. The dominance x dominance (l) interaction was larger than the additive x additive (i) and additive x dominance (j) effects put together, while for the main effects the dominance component (h) was greater than the additive (d) component. The dominance (h) and domi-nance x domidomi-nance (l) effects were in the opposite direc-tion, suggesting that duplicate-type epistasis occurred in most cases and indicating predominantly dispersed alleles at the interacting loci (Jinks and Jones 1958). Dominance gene effects were found to be relatively more important, as indicated by the fact that in all cases thedominance(h )val-ues were higher than the additive (d) values.
In the case of the plant height trait the additive x dom-inance (j) and dominance x dominance (l) effects were sig-nificant in the VS 9510 x Co 1 cross, while the additive x dominance (j) effect was significant only in the Cianno 13/10 x VRI 1 cross. This trait also showed duplicate-type epistasis in all cases. The dominance x dominance (l)
non-allelic interaction was higher than either the dominance (h) or additive (d) effects. Although additive (d) and additive x additive (i) gene effects were significant for the ‘plant height’ trait non-additive gene effects appear to have been more important. Ganesh (1999) reported duplicate epistasis for this trait.
Table 1- Mean and their standard error of six generations with four crosses for seven traits.
Generation and trait VS9501 X CO1 NIC7907 X TMV3 Cianno 13/10 x VRI 1 Si 1115/1 x TMV 3 Days to maturity
P1 P2 F1 F2 B1 B2
99.90±1.33 102.2±0.81 102.46±0.27 103.3±0.26 101.25±0.25 102.23±0.35
77.66±0.61 90.69±1.08 102.47±0.44 100.48±0.44 99.32±0.42 102.24±0.36
105.33±0.80 102.26±0.81 103.46±0.25 103.44±0.26 101.25±0.25 102.24±0.35
93.19±1.48 90.69±1.08 107.42±0.29 102.42±0.29 100.81±0.27 102.24±0.35 Plant height (cm)
P1 P2 F1 F2 B1 B2
117.33±1.77 109.26±1.81 128.20±1.60 112.98±2.65 116.60±2.01 87.73±2.78
100.93±1.83 108.06±3.21 118.53±2.98 104.91±1.79 115.66±1.78 111.50±1.76
91.60±1.22 97.06±1.80 99.46±1.91 102.35±3.21 101.93±1.83 96.90±2.7
95.56±2.02 94.36±2.21 131.00±2.98 113.50±1.96 105.23±1.79 110.30±1.61 Branches per plant
P1 P2 F1 F2 B1 B2
4.86±0.30 6.00±0.22 6.80±0.17 5.10±0.14 3.70±0.22 4.50±0.17
4.00±0.30 4.33±0.17 5.00±0.24 4.13±0.17 3.83±0.21 3.47±0.26
3.80±0.03 3.90±0.02 5.30±0.06 5.50±0.06 5.26±0.10 4.40±0.07
2.60±0.30 2.86±0.22 4.06±0.20 2.43±0.17 2.30±0.06 3.13±0.28 Capsules per plant
P1 P2 F1 F2 B1 B2
43.20±1.9 104.03±6.16 102.19±8.63 78.87±2.45 53.97±1.79 67.62±2.68
41.83±3.59 58.93±3.50 101.26±4.02 87.66±3.15 73.72±2.29 64.57±3.32
98.73±1.17 83.26± 1.30 106.63 ±2.7
74.01±1.9 67.20±2.48 53.75±2.00
31.73±3.49 50.70±4.02 133.86±7.6
67.09±2.2 43.22±1.6 43.49±1.44 Capsule length (cm)
P1 P2 F1 F2 B1 B2
2.33± 0.01
2.34±0.02 2.42±0.01 2.40±0.02 2.38±0.05 2.49±0.05
3.09±0.01 2.32±0.04 2.94±0.02 2.81±0.09 3.04±0.004 2.65±0.06
1.84±0.01 2.08±0.01 2.25±0.02 2.10±0.01 2.27±0.04 2.23±0.04
2.30±0.01 2.34±0.01 2.42±0.01 2.40±0.06 2.38±0.04 2.49±0.04 Weight of 103seeds (g)
P1 P2 F1 F2 B1 B2
2.64±0.05 3.39±0.04 3.92±0.06 3.77±1.33 3.10±0.05 3.77±0.06
2.61±0.10 3.22±0.08 3.86±0.14 3.34±0.06 3.77±0.06 3.89±0.05
2.70±0.09 3.34±0.06 3.72±0.04 3.63±0.05 3.75±0.04 3.59±0.08
2.20±0.09 3.34±0.09 3.47±0.10 3.72±0.04 3.11±0.05 3.93±0.06 Seed yield per plant (g)
P1 P2 F1 F2 B1 B2
20.90±0.32 25.57±0.23 40.68±0.41 25.29±0.39 19.72±0.31 24.21±0.48
20.10±0.26 21.01±0.28 32.12±0.26 25.03±0.28 26.17±1.5 25.08±0.41
11.53±0.28 23.17±0.26 30.13±0.48 18.04±0.77 14.61±1.01 18.05±1.07
For the ‘number of branches per plant’ trait thel ef-fect was found to be the most important in the VS 9510 x Co 1 and NIC 7907 x TMV 3 crosses while for the Cianno 13/10 x VRI 1 cross both additive x dominance (j) and dom-inance x domdom-inance (l) effects were significant. Dupli-cate-type epistasis was complementary in the Si 1115/1 x TMV 3 cross. Duplicate type of epistasis was also reported for this trait by Kumaret al.(1998).
In regard to the ‘number of capsules per plant’ trait dominance gene effects were found to be relatively more
important because of the dominance (h) values were higher than the additive (d) in three out of four cases and genes with negative effects were dominant over genes with posi-tive effects. Non-allelic addiposi-tive x dominance (j) and domi-nance x domidomi-nance (l) interactions and duplicate epistasis were observed for this trait. Additive gene action (d) along with an dominance x dominance (l) type interaction for this trait were found in the Cianno 13/10 x VRI 1 cross.
For the ‘seed yield per plant’ trait all the gene interac-tion effects were non-significant for the NIC 7907 x TMV 3
Table 2- Means ± standard error and scaling test and genetic effects for different sesame traits.
Cross A B C m$ [d$] [h$] [i$] [$j] [$l] Type of Epistasis Days to maturity
C1 - - * 106.29* ± 1.59 -1.18 ± 1.78 -10.41* ± 4.41 -5.84* ± 1.38 0.20 ± 0.89 5.99 ± 3.30 D C2 * * * 82.72* ± 2.17 6.71* ± 0.62 51.27* ± 5.19 1.21 ± 2.08 3.85* + 0.83 -31.53* ± 3.71 D C3 * - - 110.48* ± 1.45 1.53* ± 0.57 -20.27* ± 3.72 -6.86 * ± 1.33 -2.52* ± 0.72 12.22* ± 3.10 D C4 * * * 95.53* ± 1.75 1.25 ± 0.92 20.63* ± 4.52 -3.58* ± 1.48 -2.67* ± 1.02 -13.70* ± 3.50 D Plant height
(cm)
C1 - * * 156.86* ± 16.76 -3.74* ± 2.35 -146.85* ± 42.41 -43.26* ± 16.63 32.60* ± 6.13 118.19* ± 26.63 D C2 - - - 69.70* ± 20.48 -3.56 ± 2.12 92.02 ± 51.78 34.80 ± 20.38 7.67 ± 7.3 -43.19 ± 32.28 D C3 * - * 108.46* ± 6.63 -2.70* ± 1.26 -9.30 ± 16.26 -14.09* ± 6.51 5.42* ± 2.37 0.31 ± 10.9 D C4 - - - 117.90* ± 6.27 -0.60 ± 0.22 -30.70 ± 0.69 -22.94 ± 1.23 -5.67 ± 0.96 43.80 ± 1.56 D Number of branches per plant
C1 * * * 9.43* ± 0.98 -0.57* ± 0.20 -14.69* ± 2. 50 -4.0*±.096 -0.23 ± 0.39 12.06* ± 1.60 D C2 * * - 6.07* ± 1.01 -0.17 ± 1.40 -6.74* ± 2.45 -1.92 ± 1.00 0.53 ± 0.33 5.67* ± 1.57 D C3 * - * 6.74* ± 0.44 -0.01 ± 0.23 -3.30* ± 1.17 -2.91* ± 0.43 0.94* + 0.19 1.89* ± 0.82 D C4 * - * 1.59 ± 0.18 -0.13 ± 0.25 0.89 ± 2.28 -1.14 ± 0.77 -0.70 ± 0.42 1.58 ± 1.59 C Number of capsules per plant
C1 * * - 145.93*±12.59 -30.41* ± 3.23 -224.51* ± 29.25 -72.31* ± 11.76 16.77* ± 4.5 180.77* ± 24.56 D C2 - * * 124.42* ± 15.17 -8.55* ± 2.51 -124.01* ± 35.72 -74.08* ± 14.95 17.69* ± 4.75 100.80* ± 22.56 D C3 * * * 145.16* ± 10.01 7.73* ± 0.90 -246.04* ± 24.67 -54.16* ± 9.96 5.72 ± 3.32 207.52*.±15.94 D C4 * * * 136.16* ± 10.10 -9.50* ± 2.66 -273.90* ± 23.50 -94.93* ± 9.80 -9.23* ± 3.44 271.6* ± 20.3 D Capsule length (cm)
C1 - - - 2.19* ± 0.26 -.005 ± 0.035 0.59 ± 0.76 0.17 ± 0.27 -0.11 ± 0.12 -0.37±0.51 D C2 - - - 2.51* ± 0.16 0.39* ± 0.03 0.61 ± 0.38 0.14 ± 0.16 0.05 ± 0.05 -0.23±0.24 D C3 * - * 1.43* ± 0.07 -0.12* ± 0.03 1.65* ± 0.17 0.54* ± 0.06 0.16* ± 003 -0.82*±0.11 D C4 - - - 2.19* ± 0.29 -0.01 ± 0.03 0.62 ± 0.76 0.14 ± 0.27 -0.10 ± 0.12 -0.37 ± 0.51 D 1000 seed weight (g)
C1 - - * 3.02* ± 0.25 0.29* ± 5.24 1.57 * ± 0.65 0.66* ± 0.24 0.08 ± 0.20 -0.14 ± 0.46 D C2 - - * 3.02* ± 0.25 -0.31* ± 0.08 1.92* ± 0.64 0.90* ± 0.24 -0.05±.22 -0.452*±0.43 D C3 - - * 2.32* ± 0.25 0.18* ± 0.07 3.45* ± 0.659 1.21* ± 0.24 0.00 + 0.21 -1.36* ± 0.45 D C4 - - * 3.38* ± 0.29 0.47* ± 0.08 0.62 ± 0.77 0.42 ± 0.28 0.12 ± 0.26 0.24 ± 0.53 C Seed yield per plant (g)
C1 * * * 36.94* ± 1.94 -2.33* ± 0.20 -49.90* ± 4.66 -13.70* ± 1.93 -2.16* ± 0.60 53.64* ± 2.90 D C2 - * * 18.18* ± 3.34 -0.45* ± 0.19 13.47 ± 9.69 2.38 ± 3.33 1.55 ± 1.58 0.47 ± 6.41 C C3 * * * 24.19* ± 4.28 -5.82* ± 0.23 -30.53* ± 10.83 -6.84 ± 4.27 2.38 ± 1.50 36.46* ± 6.70 D C4 * * - 26.24* ± 1.60 -3.10* ± 1.90 -10.73* ± 4.07 -8.33* ± 1.59 -1.15* ± 0.59 16.61* ± 2.58 D
cross, while the remaining three crosses showed type domi-nance x domidomi-nance (l) effects. These results indicate that seed yield per plant is predominantly controlled by domi-nance x domidomi-nance (l) type interaction effects. The domi-nance (h) and dominance x dominance (l) gene effects showed opposite signs, indicating the presence of duplicate dominant epistasis in the expression of this trait.
A additive x dominance (j) type interaction was ob-served for the ‘number of capsules per plant’ and ‘seed yield per plant’ traits and thehgene effect was found to be predominant in the expression of these traits. These find-ings are in good agreement with those of Deenamani and Dorairaj (1994) for the ‘number of capsules per plant’ trait and with Ganesh (1999) for the ‘seed yield per plant’ trait, who also reported duplicate epistasis for this trait. Only the Cianno 13/10 x VRI 1 cross showedd,iandjeffects for capsule length, a simple additive-dominance model being adequate for the other three crosses To have a positive shift in the expression of the phenotypic mean it is essential to harness both the additive and non-additive gene effects prevalent in the ‘capsule length’ strength.
The ‘weight per 1000 seeds’ trait was additive and complementary for the Si 1115/1 x TMV 3 cross while ad-ditive (d), dominance x dominance (l) and additive x domi-nance (j)effects and duplicate epistasis were important in the three other crosses. Thed,hand additive x additive (i) type gene interactions indicate that this trait is under the control of both fixable and non-fixable gene effects. The additive x additive (i) type gene interaction and duplicate epistasis seen in this trait suggest the possibilities of obtain-ing transgressive segregants in later generations.
The additive effects and gene interaction dominance x dominance (l) or other type digenic complementry gene interaction can be exploited effectively by selection for the improvement the characters. Use of reciprocal recurrent se-lection or Bi-parental mating suggested to improve the characters when both additive and non- additive gene ef-fects are involved in the expression of these traits. Presence of non-additive gene for days to maturity, number of branches per plant, number of seed capsule per plant and seed yield per plant indicating that conventional selection procedure may not be effective enough for improvement of yield. Therefore postponement of selection in later genera-tions or intermating among the selected segregants
fol-lowed by one or two generations of selfing could be suggested to break the undesirable linkage and allow the accumulation of favorable alleles for the improvement of this trait.
The different types of gene effects estimated provided a test for gene action and are useful for analyzing the ge-netic architecture of a crop so as to further improve desir-able traits. The estimates obtained from each cross may be unique to that cross and may not be applicable to the paren-tal population. Additive genetic variance formed the major part of the genetic variance for the important yield compo-nent ‘weight per 1000 seeds’ trait. Therefore genetic im-provement in the ‘seed yield per plant’ trait would be easier through indirect selection for a component trait such as the ‘weight per 1000 seeds’ trait than through direct selection for seed yield itself.
References
Deenamani IE and Stephen Dorairaj M (1994) Genetics of quanti-tative characters associated with capsules in Sesamum indicumL. Madras Agric J 81:241-243.
Ganesh SK and Sakila M (1999) Generation mean analysis in ses-ame (Sesamum indicumL.) crosses. Sesame and Safflower Newsl 14:8-14.
Jinks JL and Jones RM (1958) Estimation of the components of heterosis. Genetics 43:223-224.
Kearsey MJ and Pooni HS (1996) The Genetic Analysis of Quan-titative Traits.1st edition. Chapman and Hall, London. Kumar ST, Thangavelu S and Sree Rangasmay SR (1998) Genetic
analysis of seed size characters in sesame. (Sesamum indicumL.). Sesame and Safflower Newsl 13:26-32. Mather K (1949) Biometrical Genetics. Mather and Co., London. Mather K and Jinks JL (1971) Biometrical Genetics: The Study of
Continuous Variations. Chapman and Hall Ltd., London, 382 pp.
Mosjidis JA (1982) The inheritance of oil content and its fatty acid composition in the sesame seed. (Sesamum indicumL.) and the study of their correlations. Diss Abstr Intermat B 42:3947B.
Pathak, HC and Dixit, SK (1988) Genetic analysis for single stemmed sesame (Sesamum indicum L.). Indian J Genet 48:325-330.
Singh RF and Chaudhary BD (1985) Biometrical Methods in Quantitative Genetic Analysis. Kalyani Publishers, New Delhi, 302 pp.