1 Recebido para publicação em 27.8.2013 e aprovado em 2.12.2013. 2 Universidade de Maringá, Maringá-PR, Brazil; <[email protected]>.
M
ULTIPLER
ESISTANCE OFConyza sumatrensis
TOC
HLORIMURON-E
THYL AND TOG
LYPHOSATE1Resistência Múltipla de
Conyza sumatrensis
ao Chlorimuron-Ethyl e ao Glyphosate
SANTOS, G.2, OLIVEIRA JR., R.S.2, CONSTANTIN, J.2,FRANCISCHINI, A. C.2, and OSIPE, J. B.2
ABSTRACT - Weed resistance to herbicides has been a major issue in Brazil, mainly due to the inefficiency of the herbicides used in no-till areas and to the high cost of these herbicide treatments. Failures in controlling the weed Conyza have been reported in Western and Northern grain crop areas in Paraná (Brazil). This work aimed to evaluate the potential occurrence of C.sumatrensis biotypes resistant to the herbicides chlorimuron-ethyl and glyphosate. Experiments were carried out under greenhouse conditions with four biotypes (Cascavel-2, Toledo-4, Tupãssi-6, and Assis Chateaubriand-7) possibly resistant to, as well as a population considered susceptible to chlorimuron-ethyl and glyphosate. To obtain dose-response curves, eight herbicide doses of chlorimuron-ethyl (0, 2.5, 5, 10, 20, 40, 80 and 160 g ha-1) and glyphosate (0, 90, 180, 360, 720, 1,440, 2,880 and 5,760 g e.a. ha-1) were applied
and weed control and shoot biomass evaluations were made. Results provided evidence that two biotypes (Cascavel-2 and Tupãssi-6) were resistant to glyphosate and four biotypes (Cascavel-2, Toledo-4, Tupãssi-6 and Assis Chateaubriand-7) were resistant to chlorimuron-ethyl. Multiple resistance to glyphosate and chlorimuron was confirmed for biotypes Cascavel-2 and Tupãssi 6. This is the first report on multiple resistance in Conyza sumatrensis, worldwide.
Keywords: EPSPs,sulfonilureas, ALS-inhibitors, Sumatran fleabane, multiple resistance.
RESUMO - A resistência de plantas daninhas aos herbicidas tem causado grandes problemas,
principalmente em relação à ineficácia dos tratamentos herbicidas utilizados em plantio direto e ao aumento do custo desses tratamentos. Falhas de controle após a aplicação de herbicidas em áreas com buva têm sido observadas nas regiões norte e oeste do Estado do Paraná, Brasil. Este trabalho
teve como objetivo avaliar a ocorrência de resistência de biótipos de Conyza sumatrensis aos
herbicidas chlorimuron-ethyl e glyphosate. Os experimentos foram conduzidos em casa de vegetação, avaliando-se quatro biótipos (Cascavel-2, Toledo-4, Tupãssi-6 e Assis Chateaubriand-7) com suspeita de resistência, além de uma população suscetível ao chlorimuron-ethyl e ao glyphosate. Para obter a curva de dose-resposta, foram utilizadas oito doses crescentes de chlorimuron-ethyl (0, 2,5, 5, 10,
20, 40, 80 e 160 g ha-1) e de glyphosate (0, 90, 180, 360, 720, 1.440, 2.880 e 5.760 g e.a. ha-1).
Foram avaliadas as porcentagens de controle e o acúmulo de massa seca da parte aérea dos biótipos. Mediante os resultados obtidos, confirmou-se a resistência de dois biótipos (Cascavel-2 e Tupãssi-6) ao glyphosate e de quatro biótipos ao chlorimuron-ethyl (Cascavel-2, Toledo-4, Tupãssi-6 e Assis Chateaubriand-7). Resistência múltipla ao glyphosate e ao chlorimuron foi confirmada nos biótipos Cascavel-2 e Tupãssi-6. Trata-se do primeiro relato de ocorrência de resistência múltipla em Conyza sumatrensis no mundo.
INTRODUCTION
It is common knowledge that the main herbicide used for weed management in crop production systems involving tillage is glyphosate. Plants that grow in that environment are continuously exposed to herbicide selection pressure, which increases the risk of resistant biotypes selection.
The weed resistance to herbicides is defined as the natural and inheritable ability of certain biotypes within a population to survive and reproduce after exposure to doses of herbicide that would be lethal to normal (susceptible) individuals of the same species (Christoffoleti and López-Ovejero, 2004). It is a natural phenomenon that occurs spontaneously in the populations, and therefore the herbicide is not the causative agent, but the selector of the resistant individuals who are at low initial frequency (López-Ovejero et al., 2006).
Less than 30 years after the diagnosis of the first case of resistance, there are currently over 300 known cases of resistant species in about 40 countries (Heap, 2012); in some cases, the biotypes show resistance to one or more mechanisms of action.
Given the finding of the genus Conyza plants resistant to glyphosate in Brazil (Santos et al., 2012), a large number of farmers began using herbicides of other mechanisms of action mixed with glyphosate in order to obtain desiccation of the areas of horseweed during the off season. The main herbicides used in this modality have been mainly ALS inhibitors (chlorimuron, metsulfuron and diclosulam) and, to a lesser extent, the PPO inhibitors (flumioxazin and sulfentrazone).
Currently there are 116 weed species described as resistant to ALS-inhibiting herbicides; in Brazil, species with resistant biotypes have already been registered. The first case of embodied resistance occurred in 1993 in Bidens pilosa plants, in the State of Mato Grosso do Sul, and three years later were identified Bidens subalternans biotypes resistant to ALS inhibitors. Among the cases of resistance to ALS inhibitors occurring in Brazil, the largest number of reports are those related to Euphorbia heterophylla.
In the case of horseweed have been identified to date five cases of C. canadensis resistant to ALS in Israel, the United States and Poland, and one case of C. bonariensis, in Israel. To date, there are no reports of biotypes of C. sumatrensis resistant to ALS-inhibiting herbicides worldwide (Heap, 2012).
One of the most adopted practices in the management of resistant weeds is the use of herbicides with different mechanisms of action; however, the choice of a different mechanism of action may also constitute a risk of selecting biotypes of weeds also resistant to the alternative herbicide if this is applied as the only control option (Gould, 1995), and thus, there are cases of multiple resistance.
The use of glyphosate and of chlorimuron-ethyl in desiccation of Conyza plants has not shown satisfactory control recently, especially in western Paraná, leading to suspicion that biotypes of this species resistant to these herbicides is occurring. Thus, there is a need to ascertain the causes of failures in the control and see if the event is linked to cases of development of multiple resistance or herbicide misuse.
The aim of this study was to evaluate the possibility of multiple resistance to chlorimuron-ethyl and glyphosate herbicides in Conyza sumatrensis biotypes from areas with a history of combined use of these herbicides.
MATERIALS AND METHODS
Seed collection and identification of plants in the State of Paraná
Horseweed seeds collection were conducted in the cities of Campo Mourão, Peabiru, Engenheiro Beltrão and Floresta, aiming at the botanical identification of biotypes in the State of Paraná and serving as a basis for further studies.
The collection of plants for the removal of the seeds was done on 2/17/2010, and 17 plants per site were collected. At the time of the collection, each plant was individually placed in a paper bag, which was subsequently placed in a forced ventilation oven at 65 oC for
The collected seeds were germinated in a greenhouse (5/2010). Daily irrigation was performed until the plants reached full bloom (1/2011), when their cutting was done, for making the unicatas.
The unicatas were sent to the Biology Institute – Herbarium Uberlandense of Universidade Federal de Uberlândia, where they were cataloged and classified as Conyza sumatrensis. Currently, samples collected for identification are available in the Herbarium of Universidade Estadual de Maringá (HUEM, 2011), under numbers HUEM 21290, HUEM 21291, HUEM 21292, HUEM 21293 and HUEM 21294.
Experiments aiming to evaluate the resistance
After the identification of the biotypes in the State of Paraná, a new collection was held in different points from the previous ones, assessing the possible resistance of C. sumatrensis to glyphosate and to chlorimuron-ethyl. The experiments were conducted in a greenhouse, in the period between May and July 2011.
The second collection of the C.sumatrensis plants to remove the seeds with suspected resistance to glyphosate was held on 1/31/ 2011, when the seeds were presented in a stage of full physiological maturity. The plants were taken from areas with a history of applications of glyphosate and chlorimuron-ethyl in weed desiccation and cultivation of RR soy of at least four years, with observations of the producers for their lack of control after the use of these herbicides.
Susceptible seeds were collected in the city of São Jorge do Ivaí, on farm “Meu cantinho”, where there was no history of use of glyphosate and chlorimuron-ethyl in the last eight years, and the removal of weeds was made by mechanical or hand weeding.
At the time of the collection, each plant was individually placed in a paper bag, which was subsequently placed in a forced ventilation oven at 45 oC for 24 hours to facilitate removal
of the seeds. The collection sites are shown in Figure 1.
The seeds of C. sumatrensis were sown in May 2011 in vessels with a capacity of 3 dm3
of soil, being placed superficially (~0.5 cm). After collecting the soil, the samples were subjected to separation of roots, clods, straw and other impurities, using a sieve with a mesh of 4 mm.
About a week after sowing, most seeds had germinated. When the plants reached two to three true leaves, a thinning was made, leaving two plants per pot.
To obtain the dose-response curve of the control C. sumatrensis compared to glyphosate and chlorimuron-ethyl were applied doses corresponding to [!, ¼, ½, 1, 2, 4 and 8 times the recommended dose (Moreira et al., 2007) for applying post-emergence, besides a control without herbicide.
Currently there is no record of glyphosate and chlorimuron-ethyl to control C.sumatrensis in Brazil. In the case of glyphosate, it was decided to use the recommended doses for C. bonariensis, since work by González-Torralva et al. (2010) showed that, among C. sumatrensis, C. bonariensis and C. canadensis, the species most susceptible to glyphosate was the first one, with values of I50 of 2.9 ± 3.1; 15.7 ± 0.8; and 34.9 ± 1.6 g i.a. ha-1
of glyphosate, respectively. Therefore, the dose of glyphosate recommended and used was of 720 g a.e. ha-1, and of chlorimuron-ethyl,
20 g ha-1 (Moreira et al., 2007; Rodrigues and
Almeida, 2011).
The doses of glyphosate (g e.a. ha-1)
assessed were of 0, 90, 180, 360, 720, 1,440, 2,880 and 5,760, and the ones of chlorimuron-ethyl (g ha-1), of 0, 2,5, 5, 10, 20, 40, 80 and
160. In the case of chlorimuron was used mineral oil at 0.05 v/v in all applications. The application was made when the plants presented five to six true leaves (1.5 ± 0.2 cm of height).
For herbicide applications, pressurized coast sprayer was used with constant pressure of 35 lb pol-2, equipped with three XR 110.02
nozzles, spaced of 0.5 m apart from each other and positioned 0.5 m from the surface of the targets, providing an application spray volume of 200 L ha-1 of spray.
Assessments and analysis of the data
Percentage assessments of control were performed (visual scale, 0-100%, where 0 means absence of symptoms and 100 the total death of the weeds) at 28 days after application (DAA), being considered acceptable control values ≥ 80% and satisfactory control ≥ 90%. At 28 DAA, the aerial part of the C.sumatrensis plants remaining was harvested by cutting
close to the ground and wrapped in paper bags, which were placed in a forced ventilation oven at 65 oC to constant weight and weighed,
so that the accumulation of dry mass could be obtained from each experimental unit (g per pot). The dry mass was corrected for percentage values by comparing the mass obtained in the herbicide treatments with the mass of the control (zero dose) considered 100%.
The data were initially subjected to analysis of variance and the F test. The data for the dose-response curves were fitted to the model of nonlinear regression of logistical type. The variable control is adjusted to the model proposed by Streibig et al. (1988):
cb
x
a
y
1
in which: y = percent control; x = dose of the herbicide (g e.a. ha-1) or (g i.a. ha-1); a, b and c
= estimated parameters of the equation so that: a = mathematical asymptote between the maximum point and the minimum point of the variable; b = dose that provides 50% of the asymptote; and c = slope of the curve around b.
From the log-logistic equations were produced dose-response curves. Based on the adjusted models, there was the calculation of the dose of herbicide that would provide 50% and 80% of control (I50 and I80). To perform the calculation, it was decided to reverse the log-logistic model, leaving it due to x, according to Carvalho et al. (2005):
c
y
a
b
x
*
1
From the I50 values, the resistance factor was obtained (FR) for each combination of biotypes suspected resistant and susceptible biotypes of each species. The resistance factor (FR = R/S) expresses the number of times that the dose necessary to control 50% of the
resistant biotypes is higher than the dose that controls 50% of the susceptible biotypes.
To characterize a biotype resistant to glyphosate and chlorimuron-ethyl, were used two concepts: the resistance factor (FR) and I80 (dose required to achieve 80% control). I. e., for the biotype to be considered resistant to glyphosate, it should necessarily present FR > 1.0 and I80 > 720 g e.a. ha-1 (recommended
dose). In the case of the chlorimuron-ethyl, should necessarily present FR > 1.0 and I80 > 20 g ha-1 (recommended dose).
RESULTS AND DISCUSSION
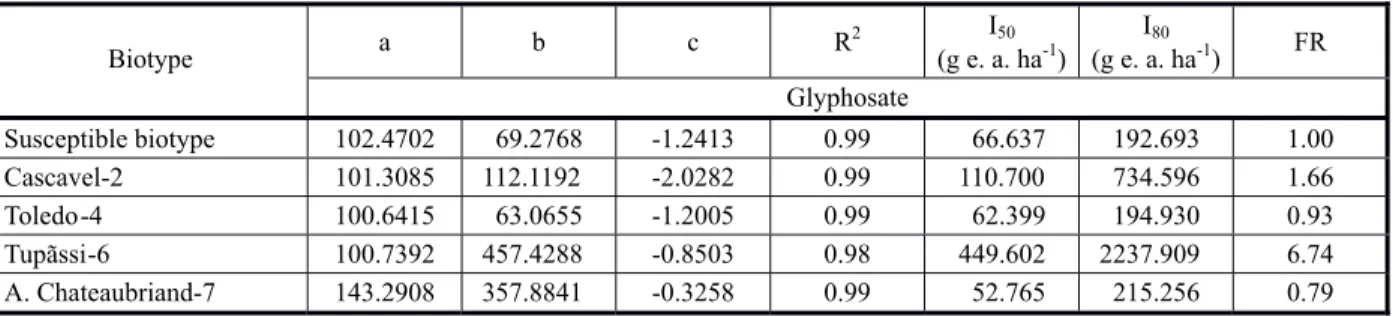
From the parameters of the log-logistic models (Table 1), were calculated the values of I50, I80 and I90, being I50 the value that characterizes the level of susceptibility of the biotypes of C.sumatrensis to the herbicide; I80, the necessary dose to obtain control of 80%; and I90, the necessary dose to obtain optimum control of the weed (90%). The resistance factor values are also presented (FR = R/S).
Two cases of resistance to glyphosate were also recorded: in biotypes Cascavel-2 (F = 1.66 and I80 =734.6 g e.a. ha-1) and
Tupãssi-6 (F = 6.74 and I80 = 2237.9 g e.a. ha-1).
It is highlighted in this case that Cascavel-2 was considered resistant, being in the threshold of the concepts used as resistant biotype (FR > 1.0 and I80 > 720 g e.a. ha-1). Both
biotypes were from the western region of Paraná, in areas with a history of applications of glyphosate and succession planting of soybean-corn, thus increasing the selection pressure of these biotypes.
Table 1 - Estimates of the parameters a, b and c and of the coefficient of determination (R2) of the logistical-log model adjusted for glyphosate, doses that provided 50 and 80% of control (I50 and I80) and factor of resistance R/S (FR), obtained for the biotypes
The control variables and dry mass (Figure 1) showed results in agreement with what has been discussed above, i.e., higher doses of applications were required on the suspected resistant biotypes to obtain the same reduction found for the susceptible biotype.
One hypothesis is that these populations of young plants of C. sumatrensis adapted and resistance occurs due to the change in the target site of EPSPS, proposed by Powles and Preston (2006). However, further studies are needed to investigate possible causes of resistance, since the mechanisms of resistance to glyphosate can be divided into two groups: those that involve changes to the destination (action site) and those which are not related to the destination site of the herbicide (Powles and Preston, 2006; Powles and Yu, 2010).
Mutations in the target location and gene amplification are examples of mechanisms of resistance in the place of herbicide action. Reduced absorption, translocation, low and sequestration of glyphosate in the vacuole are examples of resistance that do not involve the target site of the herbicide (Powles and Shaner, 2001; Powles and Yu, 2010).
According to the control data and percentage of dry mass (Figure 2) and the parameters of the log-logistic models adjusted for the controls obtained with the application of growing doses of chlorimuron-ethyl on the biotypes of C. sumatrensis, the resistance of the populations Cascavel-2, Toledo-4, Tupãssi-6 and Assis Chateaubriand-7 can be confirmed. The resistance factors (FR) for
Figure 2 - Dose-response curve: (A) % of control and (B) % dry mass from the plants of C. sumatrensis of the biotypes
cascavel-2, Toledo-4, Tupãssi-6 and Assis Chateaubriand-7, subjected to increasing doses of chlorimuron-ethyl.
these cases varied between 18.93 and 47.02 and the doses of I80 varied from 44.68 to 232 g ha-1 (Table 2).
Table 2 - Estimates of the parameters a, b and c and of the coefficient of determination (R2) of the logistical-log model adjusted for chlorimuron-ethyl, doses that provided 50 and 80% of control (I50 and I80) and factor of resistance R/S (FR), obtained for the biotypes of Conyza sumatrensis at 28 days after application (DAA). Maringá, PR – 2011
These areas have a history of application of chlorimuron-ethyl associated with glyphosate on weed control. In the case of the area of Tupãssi-6, the owner opted to plant STS soy (cultivars tolerant to herbicides of the sulphonylurea group) in harvest 2010/ 11. When collecting the plants of Conyza to remove the seeds, it was observed in this area a high population of horseweed plants, which was not expected by the producer nor by the professionals responsible for the area.
Note that the resistance factors (FR) found for chlorimuron-ethyl are larger than those found for glyphosate. This demonstrates that, in the case of this herbicide, the values of FR show a magnitude of 4-50 times higher than those observed for the glyphosate.
In the case of ALS herbicide inhibitors, reducing the sensitivity of the active site caused by points of mutations in the chain of the acetolactate synthase enzyme gene (ALS) is the most common causative mechanism of resistance of weeds (Mallory-Smith & Namuth, 2012), which causes high values of resistance factors (FR).
In Brazil, were already identified 11 species resistant to ALS (Heap, 2012). The resistance of spp. to the ALS inhibitors herbicides has been studied for some years. In Spain were identified biotypes of C. albida (also known as C. sumatrensis) resistant to imazapyr (Osuna and Prado, 2003). Populations of C. canadensis resistant to the ALS inhibitors herbicides were also identified in Ohio (USA), with resistance factors (FR) to cloransulam-methyl of 34 to 943 (Trainer et al., 2005).
The ALS enzyme in the resistant species can present different mutations, among them the replacement, in the A active center of ALS, of proline 197 by an alanine, glutamine, histidine, serine or threonine (Zheng et al., 2005). A new mutation was found in position 376 and the replacement of glutanina by aspartate was noted, which occurs within a conserved region of five amino acids in Amaranthus hybridus (Whaley et al., 2007), Kochia scoparia (Warwick et al., 2008), Amaranthus powellii (Ashigh et al., 2009) and Vaginalis monochoria (Imaizumi et al., 2008). This type of mutation is rare compared to that which occurs at position 197.
In general, replacements in positions Ala122 or Ser653 give little or no resistance to herbicides from the group of sulfonylureas and a high rate of resistance to herbicides of the imidazolinone group (Patzoldt et al., 2001). Likewise, replacements in Pro197 give resistance to sulfonylureas (Guttieri et al., 1992), but low or no resistance to the imidazolinone group (Tranel et al., 2012). Replacements in Trp574 or Ala205 provide resistance to the four chemical groups belonging to ALS inhibitors (sulfonylureas, triazolopyrimidines, imidazolinone and pyrimidil-benzoates). Nonetheless, when the mutation occurs in position Ala205, the levels of resistance are lower than the ones presented when the mutation occurs in position Trp574 the amino acid sequence of the DNA of the synthase acetolactate enzyme (Whaley et al., 2007).
It is possible that the cases of resistance of the biotypes of C. sumatrensis identified in this work have been due to one of the three aforementioned substitutions: Pro197, Trp574 or Ala205. Further studies should be performed, for the identification of possible mutations in the gene sequence of horseweed resistant ALS inhibitors herbicides would allow, for example, the development of molecular tests for rapid detection of resistance. Furthermore, studies are needed to obtain information on the cross-resistance of these biotypes to other ALS inhibitors herbicides.
Thus, this study has confirmed the resistance of biotypes of Conyza sumatrensis to glyphosate and to chlorimuron-ethyl, being the biotypes collected in Cascavel-2 and Tupãssi-6 resistant to glyphosate. As for chlorimuron-ethyl, the resistance of four biotypes was noticed: Cascavel-2, Toledo-4, Tupãssi-6 and Assis-Chateaubriand-7. These results suggest that the biotypes belonging to Cascavel-2 and Tupãssi-6 exhibit multiple resistance to glyphosate and chlorimuron-ethyl herbicides.
LITERATURE CITED
CARVALHO, S. J. P. et al. Curvas de dose-resposta para avaliação do controle de fluxos de emergência de plantas daninhas pelo herbicida imazapic. Planta Daninha, v. 23,
n. 3, p. 535-542, 2005.
CHRISTOFFOLETI, P. J.; LOPÉZ-OVEJERO, R. F. Definições e situação da resistência de plantas daninhas aos herbicidas no Brasil e no mundo. In: CHRISTOFFOLETI, P. J. Aspectos de resistência de plantas daninhas a
herbicidas. 2. ed. Campinas: HRAC-BR., 2004. p. 3-22.
GONZALEZ-TORRALVA, F. et al. Differential
susceptibility to glyphosate among the Conyza weed species
in Spain. J. Agric. Food Chem., v. 58, n. 7, p. 4361-4366, 2010.
GOULD, F. Comparisons between resistance management strategies for insects and weeds. Weed Technol., v. 9, n. 1, p. 830-839, 1995.
GUTTIERI, M. J. et al. DNA sequence variation in domain A of the acetolactate synthase gene of herbicide resistant and susceptible weed biotypes. Weed Sci., v. 40, n. 1, p.
670-676, 1992.
HEAP, I. International Survey of Herbicide Resistant. Disponível em: <http://www.weedscience.org/In.asp>. Acesso em: 5 jan. 2012.
HUEM, HERBÁRIO VIRTUAL DA FLORA E DOS FUNGOS. Disponível em: <http://inct.splink.org.br>. Acesso em: 25 nov. 2011.
IMAIZUMI, T. et al. Genetic diversity of sulfonylurea-resistant and -susceptible Monochoria vaginalis populations
in Japan. Weed Res., v. 48, n. 1, p. 187-196, 2008.
LÓPEZ-OVEJERO, R. F. et al. Suscetibilidade comparativa a herbicidas pós-emergentes de biótipos de Digitaria
ciliaris resistente e suscetível aos inibidores da
ACCase. Planta Daninha, v. 24, n. 4, p. 789-796, 2006.
MALLORY-SMITH, C.; NAMUTH, D. Overview and objectives of herbicide resistance: mechanisms; inheritance and molecular genetics. Disponível em: <http://plantandsoil.unl.edu>. Acesso em: 10 jul. 2012.
MOREIRA, M. S. et al. Resistência de Conyza canadensis e
C. bonariensis ao herbicida glyphosate. Planta Daninha,
v. 25, n. 1, p. 157-164, 2007.
OSUNA, M. D.; PRADO, R. Conyza albida: a new biotype
with ALS inhibitor resistance. Weed Res., v. 43, n. 1, p. 221-226, 2003.
PATZOLDT, W. L.; TRANEL, P. J. ALS mutations conferring herbicide resistance in waterhemp. Proc. North Central Weed Sci. Soc., v. 56, n. 67, p. 83-90, 2001.
POWLES, S. B.; PRESTON, C. Evolved glyphosate resistance in plants: biochemical and genetic basis of resistance. Weed Technol., v. 20, n. 3, p. 282-289, 2006.
POWLES, S. B.; YU, Q. Evolution in action: plants resistant to herbicides. An. Rev. Plant Biol., v. 61, n. 1, p. 317-347, 2010.
POWLES, S. B.; SHANER, D. L. Herbicide resistance and world grains. Boca Raton: CRC Press, 2001. 308 p.
RODRIGUES, B. N.; ALMEIDA, F. S. Guia de herbicidas.
6.ed. Londrina: Edição dos Autores, 2011. 700 p.
SANTOS, G. et al. Resistência múltipla ao glyphosate e ao chlorimuron-ethyl em biótipos de Conyza sumatrensis.
Dissertação. Disponível em: <http://www.pga.uem.br/
teses_p.php>. Acesso em: 12 jul. 2012.
STREIBIG, J. Herbicide bioassay. Weed Res., v. 28, n. 1, p. 479-484, 1988.
TRANEL, P. J. et al. ALSmutations from herbicide-resistant weeds. Disponível em: <http://
www.weedscience.org/mutations/>. Acesso em: 5 jan. 2012.
TRAINER, G. D. et al. Response of horseweed biotypes to foliar applications of cloransulam-methyl and glyphosate.
Weed Technol., v. 19, n. 2, p. 231-236, 2005.
WHALEY, C. M.; WILSON, H. P.; WESTWOOD, J. H. A new mutation in plant ALS confers resistance to five classes of ALS-inhibiting herbicides. Weed Sci.,v. 55, n. 1, p. 83-90, 2007.
WARWICK, S. I. et al. Acetolactate synthase target-site mutations and single nucleotide polymorphism genotyping in ALS-resistant kochia (Kochia scoparia). Weed Sci., v. 56,
n. 1, p. 797-806, 2008.
ZHENG, D.; PATZOLDT, W. L.; TRANEL, P. J. Association os the W574L ALS substitution with resistance to
cloransulam and imazamox in common ragweed (Ambrosia