ContentslistsavailableatScienceDirect
Toxicology
Reports
jo u r n al hom e p ag e :w w w . e l s e v i e r . c o m / l o c a t e / t o x r e p
The
renal
effects
and
initial
characterization
of
venom
from
Philodryas
nattereri
Steindachner,
1870
Marinetes
Dantas
de
Aquino
Nery
a,∗,
Natacha
Teresa
Queiroz
Alves
b,
Renata
de
Souza
Alves
c,
Daniel
Freire
de
Sousa
b,
Dalgimar
Beserra
de
Menezes
d,
Erik
de
Aquino
Nery
e,
Hermano
Damasceno
de
Aquino
f,
Rayane
de
Tasso
Moreira
Ribeiro
g,
Helena
Serra
Azul
Monteiro
haDepartmentofPhysiologyandPharmacology,DepartmentofBiology,FederalUniversityofCeará,Fortaleza,Ceará,Brazil
bDepartmentofPhysiologyandPharmacology,FederalUniversityofCeará,Fortaleza,Ceará,Brazil
cFacultyofPharmacy,FederalUniversityofCeará,Fortaleza,Ceará,Brazil
dDepartmentofPathology,FederalUniversityofCeará,Fortaleza,Ceará,Brazil
eGeneralHospitalofFortaleza,Fortaleza,Ceará,Brazil
fDepartmentofBiology,FederalUniversityofCeará,Fortaleza,Ceará,Brazil
gInstituteofBiosciences,UniversityofSãoPaulo,SãoPaulo,SãoPaulo,Brazil
hDepartmentofPhysiologyandPharmacology,FacultyofMedicine,FederalUniversityofCeará,Fortaleza,Ceará,Brazil
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received28June2014
Receivedinrevisedform4September2014 Accepted9September2014
Availableonline2October2014
Keywords:
Philodryasnattereri
Proteincontent Renalfailure Cytotoxicactivity
Chemicalcompoundsstudiedinthisarticle:
Beta-mercaptoethanol(PubChemCID: 1567)
Serumalbumin(PubChemCID:16132) MMTtetrazolium(PubChemCID:64965) Trishydrochloride(PubChemCID93573) 2-Propenamide(PubChemCID:6579) Saline(PubChemCID5234)
a
b
s
t
r
a
c
t
ThevenomofthesnakePhilodryasnattereriisamixtureofproteinsandtoxicpeptideswith severalimportantlocalandsystemicactions,whicharesimilartothoseoccurringin Both-ropssnakebites.Themechanismsinvolvedinthelocalandsystemicactionsofthisvenom areunknown.TheaimsoftheworkweretoinitialcharacterizationofP.nattererivenom andinvestigatetheeffectsofthepoisonintherenalperfusionsystemandinculturedrenal tubularcellsofthetypeMDCK(Madin–Darbycaninekidney).TheP.nattererivenomis composedmajorityofproteins(86.3%)andthispoisonpromotedchangesinallthe evalu-atedrenalparameters,mainlydecreasingrenalperfusionpressure(PP)andrenalvascular resistance(RVR)andincreasingurineflow(UF)andglomerularfiltrationrate(GFR).The mostrelevantresultwasthatthisvenomwashighlydetrimentaltotherenaltubules inde-pendentofthePPreduction,whichwasshownbyadecreaseinsodium(Na+),potassium (K+)andchloride(Cl−)electrolytetransportinthestudiedconcentrations.Theglomeruli andtubulescontainproteinbodiesandbloodextravasation,whichwereobservedby his-tologicalanalysis.ThevenomofP.nattererireducedviabilityoftheMDCKcellsonlyathigh concentrations(50and100g/mL)withanIC50of169.5g/mL.
©2014TheAuthors.PublishedbyElsevierIrelandLtd.Thisisanopenaccessarticleunder theCCBY-NC-NDlicense(http://creativecommons.org/licenses/by-nc-nd/3.0/).
∗ Firstandcorrespondingauthor.Tel.:+558533669834.
E-mailaddress:[email protected](M.D.d.A.Nery).
1. Introduction
Philodryas nattereri Steindachner, 1870 of the family Dipsadidae[1],commonlycalledthebrownracer,hasan olivegreencolorationwiththefinalportionofitsbody col-oredbrown.Thissnakeis1.20–1.60mlong,haslargeeyes
http://dx.doi.org/10.1016/j.toxrep.2014.09.005
withroundpupils,isfastandhasanintensedailyactivity (VittandColli[27]).Thesnakes’habitatisrelatedtothe environment’sphysicalstructure,foodavailability, pres-enceofpredatorsandthephysiologyofthesesnakes,which arediurnal,arborealandsemi-arboreal.Thesesnakesfeed onsmallmammals,birdsandlizards[25],areoviparous andlayfrom6to20eggs.Thesnakes’dentitionis opistho-glyphousandconnectedtotheDuvernoy’sgland.
P.nattereriisdistributedinaridandsemiaridregionsof SouthAmericaandismostcommoninnortheasternBrazil (CearáandRioGrandedoNorte).
Duringevolution,snakeshavespecializedinaffecting thevitalfunctionsoftheirpreybyreleasingalarge num-ber oftoxins (enzymes, proteinsand peptides) through venom that destabilize the physiological levels of hor-mones, alter the activity of enzymes, receptors or ion channels,andpromotecardiovascularandnervoussystem imbalanceintheirprey.Theuseofsnaketoxinsas phar-macologicaltoolsandprototypesfordrugdevelopmentis increasing[2].
Itisalsoimportanttoemphasize thattheseverityof symptoms after poisoning is related to the amount of venom inoculated, which depends on the snake’s size, ageandtime itwasfed.Poisoningcausedbyspecies of
Philodryasischaracterizedbylocalsymptomssuchaspain, swelling, erythema, bruising, renal failure and regional lymphadenopathywithnormalcoagulation[3].
The pathogenesis of the renal alterations following envenomationby Philodryasspecies isnot welldefined. Thus, this study aimed to evaluate the renal effects of
P.nattererivenom ina perfusionsystemusingdifferent concentrations of venom, to characterize possible his-tological alterations promotedbyvenom in isolated rat kidneysand tostudyvenom-inducedchangesinculture ofMadin–Darbycaninekidney(MDCK).
2. Materialandmethods
2.1. Animalsandvenomextraction
P.nattererisnakeswerecapturedonAroeirasFarminthe municipality of Upanema(5◦38′32′′Sand 37◦15′27′′W), stateofRioGrandedoNorte andtransportedtoNUROF (OphiologyRegionalNucleusofCeará).
Theanimalsweremaintainedinindividualcageswith freeaccesstowaterandfedwith15gmiceevery30days.
Venompools weremadefrommore than40 individual
snakesandcollectedfromthevenomglandintocapillary tubestopreventcontaminationwithsaliva.Afterthe out-flowofthevenomintothecapillarytube,thevenomwas frozenandlyophilized.
2.2. Analysisofproteincontent
Thevenom ofP.nattereri(1mg)waslyophilizedand resuspendedin1mLofsalinesolution.Analiquotofvenom (100L)wastakenforthequantificationofproteinsbythe methodofBradford[4]usingBio-Radreagentsandbovine serumalbumin(BSA)asastandard.
2.3. Sodiumdodecylsulphate-polyacrylamidegel electrophoresis(SDS-PAGE)
Sodium dodecyl sulphate-polyacrylamide gel elec-trophoresis(SDS-PAGE)wasperformedaccordingtothe methodof Laemmli[5].SDS-PAGE wascarried outin a 2mmverticalslabgel(10cm×8cm)consistingof stack-inggel mix, 5%total acrylamide, andmain running gel mix, 17.5% acrylamide, prepared in 3.0M Tris-HCl, pH 8.8.Samples(20g)weredissolvedinTris–HCl0.0625M, pH 6.8, containing 1% SDS and 1% -mercaptoethanol andincubated at100◦Cfor 10min.Electrophoresis was carriedoutat20mAperplatefor1.5hours.Protein mark-ers employed were myosin (212kDa), -galactosidase (116kDa),phosphorylaseB(97.4kDa),BSA(66.2kDa),egg albumin(45kDa),carbonicanhydrase (31kDa),soybean trypsininhibitor(21.4/19.7kDa)andlysozyme(14.2kDa) (AMRESCOInc.,Ohio,USA)[6,28].
2.4. Nuclearmagneticresonancespectroscopy(NMR)
For1HNMRanalysis,spectrawererecordedonaBruker
DRX-300MHzFTNMRspectrometer.VenomsamplesofP. nattereri(5mg/mL)werelyophilizedandpreparedusing dimethylsulphoxide(DMSO)asasolvent.Thespectrawere obtainedat85◦Cusingarelaxationdelayof1sandapulse widthof90◦toreachtheconditionsofquantitative anal-ysis,accordingwithAhmadetal.[7].SilicaGel60(Merck, 70–230mesh)wasusedforanalyticalTLC.Column chro-matographieswereperformedoversilicagel(Merck,60 F254230–400mesh).
2.5. Kidneyperfusion
AdultmaleWistarrats(260–320g)werefastedfor24h withfreeaccesstowater.Theratswereanesthetizedwith sodiumpentobarbitone(50mg/kg, i.p.)andaftercareful dissectionoftherightkidney;therightrenalarterywas cannulatedviathemesentericarterywithoutinterrupting thebloodflowasdescribedbyBowman[8].
The perfusion fluid consisted of modified Krebs– Henseleit solution (MKHS) of the following composi-tion (in mmol/L): 114.00 NaCl, 4.96 KCl, 1.24 KH2PO4,
0.5 MgSO4·7H2O, 2.10 CaCl2 and24.99 NaHCO3.Bovine
serumalbumin(BSA6g%;fractionV),urea(0.075g),inulin (0.075g) and glucose (0.15g) were added to the solu-tion,resultinginafinalperfusatevolumeof100mL.The pHwas adjusted to7.4. In each experiment, 100mLof MKHSwererecirculatedfor120min.Theperfusion pres-sure(PP)wasmeasuredatthetip ofthestainless steel cannula in the renal artery.Samples of urine and per-fusionfluidwerecollectedat10minintervalsforanalysisof thesodium,potassiumandchloridelevelsbyion-selective electrodes(Rapidchem744,BayerDiagnostic,UK);inulin, asdescribedbyWalseretal.[9]andmodifiedbyFonteles etal.[10];andosmolality,whichwasmeasuredinvapor pressureosmometer(Wescor5100C,USA).ThevenomofP. nattereri(10mg/mL)wasaddedtothesystem30minafter thebeginningofeachperfusion.
thepercentageofsodium(%TNa+),potassium(%TK+)and
chloride(%TCl−)tubulartransportweredetermined[11]. Theresultswerecomparedtothecontrolgroup,at30min earlyineachexperiment(n=6).Theexperimental proce-dureswereconductedaccordingtoguidelinesforthecare anduseoflaboratoryanimalsasapprovedbytheEthical Committee(68/08)fromFederalUniversityofCeará(UFC).
2.6. Renalhistologicalevaluation
Aftertherenalperfusionexperiment, both rightand left kidneys were removed and fixed in 10% formalde-hyde for histological processing. Kidney tissue was embedded in paraffin, cut into 5m sections, stained withhematoxylin–eosinand furtherprocessed for light microscopy(OlympusBX41,USA).Thephotomicrographs weretakenbymeansofadigitalcamera(NikonCoolpix 885,Japan).
2.7. Cellculture
EpithelialMadin–DarbyCanineKidney(MDCK)was cul-tivatedinRPMI1640medium(MEM)supplementedwith 10%fetalbovineserum,1%penicillin(10000IU/mL)and streptomycin(10mg/mL).Foreachexperiment,cellswere removed and incubated with trypsin-EDTA (0.25/0.02%, v/v)at37◦Catabout5min.
Afterthis,thecellswerecountedinaNeubauerchamber andsuspendedinculturemedium(1×105cells)and24h
laterusedfortheexperiments.
2.8. Cytotoxicassay
CellviabilitywasassessedbyMTT (4,5-dimetilazil-2-il)-2,5diphenyltetrazolium)assayasdescribedbyMosmann
[12]. The MDCK cells are plated in 96-well plates at a densityof105cellsandtreatedwithdifferent
concentra-tionsofP.nattererivenom(1.56,3.12,6.25,12.5,25,50, 100g/mL).After24hoftreatment,thecellswere incu-batedwith0.5mgofMTT/mLfor4h.Theformazancrystals thatresultedfromMTTreductionweredissolvedbyadding SDS(10%)toeach wellfollowedby incubationfor 17h. Theabsorbancewasreadat570nminamicroplatereader, andcellviabilitywascalculatedbycomparingthe result-ingabsorbanceswiththemeanabsorbanceofthecontrol wells(withoutvenom,consideredtobe100%viable).
2.9. Statisticalanalysis
Theresultswereexpressedasmeans±SEM(n=6). Sta-tisticalevaluationwasdeterminedbyanalysisofvariance (ANOVA)andcorrectedbytheBonferrnitest.Statistical sig-nificancewassetat5%.Theprogramsusedtoperformthe statisticalanalysiswereMicrosoftExcel2007and Graph-PadPrism5.0.
3. Results
3.1. Proteincontentofvenom
Inthepresentstudy,thevalueoftotalproteinofthe venom from P.nattereri wasof 863.9g/mg of venom,
Fig.1.ElectrophoreticprofilesofP.nattereri(P.n.)andB.jararaca(B.j.) venoms.Eachlanewasloadedwith20Lofvenom.Proteinsweresilver stained.
corresponding in 86.3% of total content of venom this species. In comparisonwithvenom ofothers species of the generaPhilodryas, suchas P.olfersii(923g/mg),P. patagoniensis(814g/mg)andP.nattereri(847g/mg)and
Bothropsjararaca(799g/mg)[13,14].ThevenomofP. nat-tereriwassimilaramongtheothersvenoms.
3.2. SDS-PAGEandNMRspectrum
SDS-polyacrylamidegelelectrophoresiswascarriedout accordingtoLaemmli[5]andshoweddistinctprotein pat-terns among the venomsof P.nattereri and B. jararaca. ThevenomofP.nattererishowedmultipleproteinbands, rangingfrom45kDato100kDa,whileinB.jararacahave proteinbandsrangingfrom45to210kDa(Fig.1).
The1HNMR spectrumof P.nattereri venomshowed
thepresence ofpeptides, amino acidssimple (phenolic, aromaticsandaliphatic)andaminoacidderivatives rep-resentedthemajorcomponentsofthisvenom(Fig.2).The peaksgeneratedcorrespondtoaminoacidsoftheprotein constituentsofthepoison,whichcomprise86.3%ofthis.
3.3. EffectsoftheP.nattererivenomintheisolatedrat kidney
Fig.2.Detailed1HNMR-spectroscopic(500MHz,CD
3OD)analysisofP.nattererivenom.(A)The1HNMRspectrumrevealingahighlycomplexcomposition withdifferentgroupsofaminoacids(B)Cozyspectrum(CD3OD)withdifferentgroupsofaminoacids.
andtothe3mg/mLreducedsignificantlyat60and90min forbothparameters,butat120minincreasesignificantly in UF, while GFRreturned tonormal perfusion(Fig.3C andD).
Na+,K+, Cl− electrolyte transport wasaltered in the perfusedkidney,regardlessofthevenomconcentration. Regardingthepercentofsodiumtubulartransport(%TNa+)
wasreducedat60,90and120minfor1and3mg/mL con-centrationsof venom and percentof potassium tubular transport(%TK+)wasreducedat60,90and120minonlyin
3mg/mLconcentration,whencomparedincontrolgroup (Fig.4AandB).
Thepercentofchloridetubulartransport(%TCl−)was reducedat90and120minfor1mg/mL,whilein3mg/mL
Fig.4. EffectsofP.nattererivenominconcentrationsof1and3mg/mLonsodium(A)potassium(B)chloride(C)tubulartransportpercentandosmotic clearance.Dataareexpressedasmean±SEMfromsixdifferentanimals.*p<0.05comparedtothecorrespondingcontrolgroupforeachinterval(ANOVA andBonferronitest).Thevenomwasaddedtothesystem30minafterthebeginningofeachperfusion.
concentrationat60, 90and 120min.Osmoticclearance showedincreaseat90and120minin1mg/mL concentra-tionofvenomofP.nattereri(Fig.4CandD).
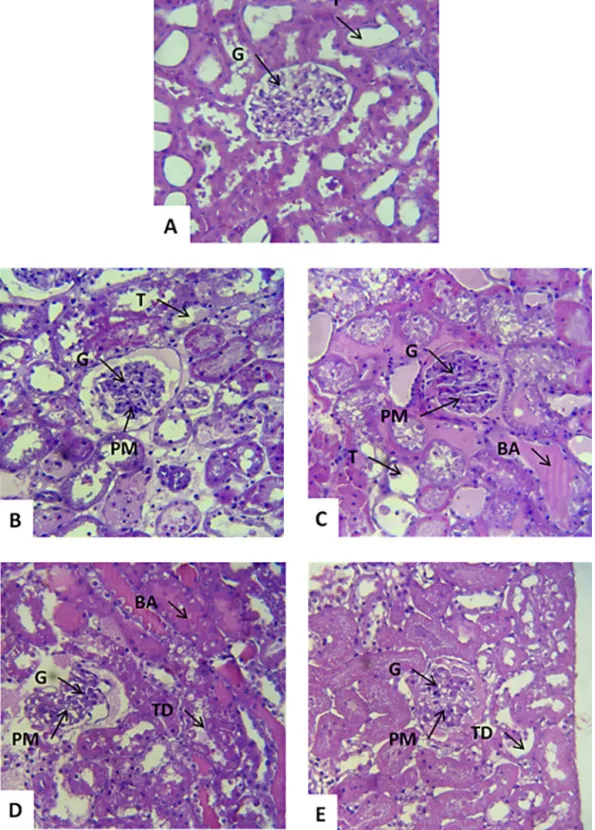
3.4. Renalhistologicalevaluation
Inthecontrolgroupcomposedofleftkidneysperfused withKrebs solution(MKHS),thekidneyshowednormal structures(glomerulus, tubule,vesselsand interstitium) (Fig.5A).
The right kidney perfused with 1mg/mL of venom
showedtubulardilation,mainlyinthedistalconvoluted tubuleandHenle’sloop,proteinaceousmaterial(PM)and bloodaccumulation(BA)withinthetubules,normalvessels andinterstices(Fig.5BandC).
Kidneysperfusedwith3mg/mLofvenomshowedan obvioustubutardegeneration,presenceofproteinaceous
material within the tubules and the Bowman spaces,
glomeruliwithslightalterationsandtubuleswith mod-eratedilatation(TD)(Fig.5DandE).
3.5. CytotoxiceffectoftheP.nattererivenomonMDCK cells
TheP.natterericytotoxicitywasassessedintherenal tubularcells(MDCKcellculture)after24hofexposureto
variousvenomconcentrations(3.125,6.25,12.5,25,50and 100g/mL).Thevenomsignificantlyreducedtheviability of theMDCK cells in the 50 and 100g/mL concentra-tionscomparedtothecontrolwithanIC50of169.5g/mL
(Fig.6).
4. Discussion
ToinitialcharacterizationofvenomfromP.nattereri,we analyzedtotalproteincontentandprofilebySDS-PAGEand NMRanalysisoflyophilizedvenom.ThepoisonofP. nat-tereriexhibited86.3%oftotalproteincontent.Thisvalueis similartothatfoundbyZelanisetal.[14]forothersspecies of thegeneraPhilodryas,suchasP.olfersii(923g/mg),
P.patagoniensis(814g/mg)andP.nattereri(847g/mg), andBothropsjararaca(Viperidaefamily)with799g/mg
[13].
RegardingSDS-PAGEanalysis,ThevenomofP.nattereri
showedalesscomplexprofileincomparisontotheother twocongenericspecies(P.olfersiiandP.patagoniensis)[14], withmajorproteinbandsrangingfrom45kDato100kDa, whileinB.jararacahavealargenumberofproteinbands rangingfrom45to210kDa.
Theseresultsmaystrengthenthehypothesesofclose similarity regardingthe actionsand activitiesof venom
Fig.6.ViabilityofMDCKcellsincubatedwithdifferentconcentrationsof
P.nattererivenomfor24h.
corroborate the eletrophoretic analysis performed by RochaandFurtado[26].
Toinvestigatetheeffectof theP.nattererivenom in thekidneywithoutinterference ofsystemic factors,we usedperfusionintheratkidney.Inthepresentstudy,we observeda decrease in perfusionpressure,renal vascu-larresistance,urinaryflowandglomerularfiltrationrate, aswell asa decrease in sodiumtransport and chloride afterthekidneywasperfusedbythevenom,whereasthe clearanceosmoticwashighercomparedwithcontrol.This agreeswithfindingsforBothropsmarajoensis[15],Bothrops insularis[24],Bothropsjararaca[16] andBothrops jarara-cussu[17].
Bothrops venom is characterized for promoting hypotensionthroughmechanismsthatpromotesystemic vasorelaxation[18].These effects are likely identical to thoseaffectingrenalPP,asweobservedinthisstudyforP. nattererivenom.
However, the reduction in almost renal parameters observedintheexperimentscannotberelatedtoa spe-cificcomponentbecausetheoverallcompositionofcrude
P.nattereri venom is unknown. Despite this limitation, mediators,suchas tumornecrosisfactor-alpha(TNF-␣), interleukin-1(IL-1),IL-6,IL-10andgammainterferon (IFN-Y),canbeinvolvedinrenalparametersdecreases(PP,GFR, RVR,TNa+,TK+andTCL−),becausevenomtypicallyinduces therelease of these substances, ashas beenpreviously reported[19].Inaddition,theremightbecompoundsthat exertdirectactiononkidneytissues.Theseinflammatory mediators,withrelaxationpotential,mayhavecontributed totheRVRreductionobservedinthisstudy.
The histological alterations promoted by P.nattereri
venomintwoconcentrationsof1 and3mg/mL, respec-tively showedinitially blood accumulation, glomerulus, tubuleswithproteinaceousmaterialanddilatationtending towarddegeneration,reversiblelesionsandthe accumula-tionofproteinsmaterialreleasedintothedamagedcells cytoplasmreflectedtheearlystagesofvenomtoxic agres-sionandthehistologicalanalysiscorroboratetheresults forthegenusBothrops[17,20,24].
ThecytotoxicpotentialofP.nattererivenomwas eval-uatedusingMDCKcells,whichisacelllinewithsimilar
morphologicalandfunctionalcharacteristicstocellsinthe distalcollectingtubulesofmammals[21].
Thresultsofcytotoxiceffectsshowedthatthevenom wastoxictocellsbecauseitsignificantlyreducedviability inconcentrationsof50and100mg/mLcomparedtothe controlwithanIC50of169.5g/mL.ThisvalueforIC50is
higherinrelationofothersspeciesofsnakes,suchas Both-ropsleucurusand Crotalusdurissuscumanensiswith1.25 and5.38g/mL,respectively[22,23].
5. Conclusions
TheP.nattererivenomwascomposedandcaused tox-icityinkidneyisolatedandinducedcelldeathoncultured MCDKcells.WedemonstratedP.nattererivenomis com-posed for proteins(86.3%) and capable ofchanging the kidneyfunctionalparameters(PP,RVR,GFR,UF,osmotic clearance, percent of sodium, potassium and chloride transport) in the isolated rat kidneys. Furthermore,the venom promotedmorphologicalalterations in therenal tubules,suchasbloodaccumulation,glomerulus,tubules withproteinaceousmaterial.Itissupposedthatthisrenal changeoccurs probablybydamagingboth vascular and glomerularsites.Regardingtheeffectsofvenominviability oftheMDCKcellswasobservedonlyathighconcentrations (50and100g/mLofvenom)withanIC50of169.5g/mL.
Thesefindingsmaybeimportantaspectsoftheprocessof toxicitymediatedbyP.nattererivenom.
Transparencydocument
TheTransparencydocumentassociatedwiththisarticle canbefoundintheonlineversion.
References
[1]H.Zaher,F.G.Grazziotin,J.E.Cadle,R.W.Murphy,J.C.Moura-Leite, S.L.Bonatto,Molecularphylogenyofadvancedsnakes(Serpentes, Caenophidia)withanemphasisonSouthAmericanXenodontines:a revisedclassificationanddescriptionsofnewtaxa,Pap.AvulsosZool. 49(2009)115–153.
[2]S.F.Paioli,CytotoxicEffectofCrotoxinonMurineMelanomaCellsand Fibroblasts(Dissertation),Inter-UnitBiotechnologyProgram, Uni-versityofSãoPaulo,SãoPaulo,2011,39pp.
[3]L.A. Ribeiro, G. Puorto, M.T.Jorge,Bites by the colubrid snake Philodryasolfersii:aclinicalandepidemiologicalstudyof43cases, Toxicon37(1999)943–948.
[4]M.M.Bradford,Arapidandsensitivemethodforthe quantifica-tionsofmicrogramquantitiesofproteinutilizingtheprincipleof protein-dyebinding,Anal.Chem.72(1976)248–254.
[5]U.K.Laemmli,Cleavageofstructuralproteinsduringtheassemblyof theheadofbacteriophageT4,Nature227(1970)680–685.
[6]S.Ahmad,H.S.Moinuddin,U.Shahab,K.Alam,A.Ali,Autoimmune responsetoAGEmodifiedhumanDNA:implicationsintype1 Dia-betesmellitus,J.Clin.Transl.Endocrinol.1(2014)66–72.
[7]S.Ahmad,S.U.Moinuddin,M.S.Khan,S.Habeeb,K.Alam,A.Ali, Glyco-oxidativedamagetohumanDNA-Neo-antigenicepitopeson DNAmoleculecouldbeapossiblereasonforautoimmuneresponse intype1diabetes,Glycobiology24(2013)281–291.
[8]R.H.Bowman,Gluconeogenesisintheisolatedperfusedratkidney, J.Biol.Chem.245(1970)1604–1612.
[9]M.Walser,D.G.Davidson,J.Orloff,Therenalclearanceofalkalistable inulin,J.Clin.Invest.34(1955)1520–1523.
[11]M. Martinez-Maldonado,R. Opava-Stitzer, Freewater clearance curvesduringsaline,mannitol,glucoseandureadiuresisintherat, J.Physiol.280(1978)487–497.
[12]T.Mosmann,Rapidcolorimetricassayforcellulargrowthand sur-vival:applicationtoproliferationandcytotoxicityassays,J.Immunol. Methods65(1983)55–63.
[13]A.C.G.PriantiJr.,W.Ribeiro,R.A.B.Lopes-Martins,R.M.Lira-Da-Silva, J.Prado-Franceschi,L.Rodrigues-Simioni,M.A.Cruz-Höfling,G.B. Leite,S.Hyslop,J.C.Cogo,EffectofBothropsleucurusvenominchick biventercervicispreparations,Toxicon41(2003)595–603.
[14]A.Zelanis,M.M.T.Rocha,M.F.D.Furtado,Preliminarybiochemical characterizationofthevenomsoffiveColubridaespeciesfromBrazil, Toxicon55(2010)666–669.
[15]I.L.Evangelista,A.M.Martins,N.R.Nascimento,A.Havt,J.S. Evan-gelista,T.B.Norões,RenalandcardiovasculareffectsofBothrops marajoensis venom and phospholipase A2, Toxicon 55 (2010) 1061–1070.
[16]H.S.A. Monteiro, M.C. Fonteles, The effect of Bothrops jararaca venomonratkidneyaftershort-termexposure:preliminaryresults, Pharmacol.Toxicol.85(1999)198–200.
[17]A.Havt,M.C.Fonteles,H.S.A.Monteiro,TherenaleffectsofBothrops jararacussuvenomandtheroleofPLA2andPAFblockers,Toxicon39 (2001)1841–1846.
[18]A.M.Soares,M.R.M.Fontes,J.R.Giglio,PhospholipasesA2myotoxins fromBothropssnakevenoms:structure–functionrelationship,Curr. Org.Chem.8(2004)1677–1690.
[19]V.L.Petricevich,C.F.P.Teixeira,D.V.Tambourgi,J.M.Gutiérrez, Incre-ment inserumcytokineandnitricoxideinmice injectedwith BothropsasperandBothrops jararacasnake venoms,Toxicon38 (2000)1253–1266.
[20]E.A.Burdmann,V.Woronik,E.B.Prado,R.C.Abdulkader,L.B. Sal-danha,O.C.Barreto,M.Marcondes,Snakebite-inducedacuterenal
failure.Anexperimentalmodel,Am.J.Trop.Med.Hyg.48(1993) 82–88.
[21]C.B.Collares-Buzato,L.S.L.DePaula,M.A.Cruz-Höfling,Impairment ofthecell-to-matrixadhesionandcytotoxicityinducedbyBothrops moojenisnakevenominculturedrenaltubularepithelia,Toxicol. Appl.Pharmacol.181(2002)124–132.
[22]T.P.Pereira,M.R.R.P.P.Bezerra,A.F.C.Torres,T.S.Brito,F.J. Batista-Lima,J.F.C.Vinhote,D.F.Sousa,R.M.Ximenes,M.H.Toyama,E.B.S. Diz-Filho,P.J.C.Magalhães,H.S.A.Monteiro,A.M.C.Martins,Renal andvasculareffectsofCrotalusdurissuscumanensisvenomandits crotoxinfraction,J.Venom.Anim.ToxinsIncl.Trop.Dis.17(2011) 333–347.
[23]I.C.De Morais,A.F.Torres,G.J.Pereira,Bothropsleucurusvenom inducesnephrotoxicityintheisolatedperfusedkidneyandcultured renaltubularepithelia,Toxicon61(2013)38–46.
[24]M.D.Braga,A.M.Martins,D.N.Amora,D.B.deMenezes,M.H.Toyama, D.O.Toyama,etal.,Purification andbiologicaleffectsofC-type lectinisolatedfromBothropsinsularisvenom,Toxicon15(2006) 859–867.
[25]FUNASA,MinistériodaSaúde.Fundac¸ãoNacionaldaSaúde.Manual deDiagnósticoeTratamentodeAcidentesporAnimaisPec¸onhentos [ManualofDiagnosisandTreatmentofSnakeBites].Brasília,October 2001.
[26]M.M.T.Rocha,M.F.D.Furtado,Caracterizac¸ãoindividualdoveneno deBothropsalternatusDuméril,Bibron &Dumérilemfunc¸ãoda distribuic¸ãogeográficanoBrasil(Serpentes,Viperidae),Rev.Bras. Zool.22(2005)383–393.
[27]L.J.Vitt,G.R.Colli,Geographical ecologyofaneotropicallizard: Ameivaameiva(Teiidae)inBrazil,Can.J.Zool.72(1994)1986–2008.