Gianluca Selvaggio
Seeking general principles in the design
of defense systems against hydrogen peroxide
Dissertation presented to obtain the PhD degree in Bioengineering
Supervisor:
Co-supervisor:
Dr. Armindo Salvador,
Assistant Researcher, Universidade de Coimbra
Dr. Manuel Nunes da Ponte,
Full Professor, Universidade Nova de Lisboa
Presidente:
Arguentes:
Vogais:
Dr. João Paulo Serejo Goulão Crespo
Dr. João António Nave Laranjinha
Dr.ª Isabel Cristina de Almeida Pereira da Rocha
Dr. Fernando José Nunes Antunes
Dr. Pedro António de Brito Tavares
Dr.ª Maria Inês Araújo Pimenta de Castro
Dr. Armindo José Alves da Silva Salvador
iii
Alla mia famiglia senza la quale non sarei mai arrivato fino a qui.
v
A human being should be able to change a diaper, plan an invasion, butcher a hog, conn a ship,
design a building, write a sonnet, balance accounts, build a wall, set a bone, comfort the dying, take
orders, give orders, cooperate, act alone, solve equations, analyse a new problem, pitch manure,
program a computer, cook a tasty meal, fight efficiently, die gallantly. Specialization is for insects.
vii
Declaration-Declaração
I declare that this dissertation is a result of my own research carried out between September 2011
and September 2016. The project was conceived and partially developed at the Computational and
System Biology Group of Dr. Armindo Salvador, Centre for Neuroscience and Cell Biology-
University of Coimbra, Portugal. Chapter 2 is an adapted version of a manuscript currently in
preparation, authored by Selvaggio G., Oliveira V., Coelho P. M. B. M. and Salvador A entitled
“Design principles for thiol redox signaling: mapping the phenotypic repertoire of the cytoplasmic 2-Cys peroxiredoxin – thioredoxin system”.
Chapter 3 was carried out at the Synthetic Biology Group under the supervision of prof. Timothy K.
Lu, Synthetic Biology Centre-MIT, Boston USA and has been published as Rubens J. R., Selvaggio
G., Lu T. K. “Synthetic mixed-signal computation in living cells.” Nat. Commun. (2016).
In vivo zebrafish experiments in Chapter 4 were performed at the Telomeres and Genome Stability Lab of Dr. Miguel Ferreira, Instituto Gulbenkian de Ciência in Oeiras, Portugal.
Declaro que esta dissertação é o resultato do meu próprio trabalho desenvolvido entre Setembro
de 2011 e Setembro de 2016. O projecto foi concebido e parcialmente desenvolvido no
Computational and System Biology Group do Dr. Armindo Salvador, Centro de Neurociências e
Biologia Celular Universidade de Coimbra, Portugal. O Capítulo 2 do
manuscrito em preparação, Selvaggio G., Oliveira V., Coelho P. M. B. M. and
Salvador A e titulado “Design principles for thiol redox signaling: mapping the phenotypic repertoire of the cytoplasmic 2-Cys peroxiredoxin –thioredoxin system”.
- é uma versão adaptada
d autoria dea
O trabalho do Capítulo 3 foi realizado no Synthetic Biology Group do prof. Timothy K. Lu, Synthetic
Biology Centre-MIT, Boston USA e foi publicado como Rubens J. R., Selvaggio G., Lu T. K.
“Synthetic mixed-signal computation in living cells.” Nat. Commun. (2016).
ix
Financial Support
–
Apoio Financeiro
This dissertation had the financial support from the FC&T doctoral fellowship SFRH/BD/51576/2011 and BIOCANT fellowship 01/2015/Sistemas Biólogicos
xi
Acknowledegments
Il primo ringraziamento va alla mia famiglia, che mi ha aiutato ad arrivare fino a qui. Senza mio
padre, mia madre e mia sorella non sarei mai riuscito a diventare l’uomo che sono adesso. In questi
trent’anni hanno fatto una marea di sacrifici per permettermi di raggiungere i miei sogni e se sono arrivato fino a qui lo devo a loro. Non passa un giorno che io non ringrazi il cielo di essere nato in
questa famiglia, e non c’é modo in cui io possa ricambiare. Tutto quello che posso dire é:” Grazie”. I have to thank many people at the end of this journey some of which became family to me, they all
help to “matar a saudade” and to retrieve happiness in moments in which was hard to find light. It
would take another thesis to thank you all for what you have done so I will try to be brief…and this
never ended good.
I was a lucky person from the very beginning when I arrived in Portugal here I meet people like
Enrico, Mariantonietta e Tina they made me call Lisbon home and feel happy in this place, I would
have not made it through the first year without they invaluable friendship. I’ve to thank also David for granting me a nick name that will haunt me for long “Fraquinho”, and Giulia made me smile in this last year. Although I was a “scary” person, and moreover an engineer, they became amazing friends.
In Coimbra I meet two of the three idiots Rui and Alessandro which made me pass beautiful days
between cigarettes and coffee, together with David which has been not only of support to me in
countless occasion but a trigger for me to get out some guts and make moves. I own to him a big
share of my happiness. Pedro joined us after a while but he has been a pillar in helping and
sustaining me in the period in the US and in all the trouble I got to get there, he was the one being
there when rushed down to FC&T to yell a bit.
These people made me call Coimbra home. I like to imagine us at the math bar in the morning, with
Ale having the n+1 coffee of the day, Rui arriving late and me David and Pedro ending up talking
about some weird double meaning until we reach that limit that is time to go away.
In Boston was a tough period and although the first memory are mice related I had the opportunity
to meet people like Mario, Carolina, Jaide and Sara. They have been special friends and one of the
funniest birthday I had, eating at a Portuguese restaurant in Boston. It has been a bless to meet
them in such a hard time.
In Oeiras, I found another major part of this big family that helped me moving forward. It was a very
hard time but thanks to the friendship and the support of Tânia and Joana I managed through it.
Our brakes with cigarettes and coffee, even if these last were not too good for my stress brought
light to those days. I was again happy to go to work because I was going to meet such amazing
people as you.
The “Telometros” lab has been a place where to be again amongst friends, Margarida and Ana the
ninja shared with me desk and bench, and “nicely” reminded me that “I was supposed to write” although they started one year before the defense to say it. Mounir has been a very good friend to
xii
Kirsten, Iris, Vanda, Akila, Rita and Raquel. You all have been so nice and kind to me that I will
never forget this past year.
To all of you my friends of the past 5 years I’m bad with words and I think I don’t have and know enough in English to say to you, with my bad accent, how much I love you, how much you represent
to me and what is the happiness you brought to my life. The only thing I can do is that to all of you
one by one I will hug you. Because is the only way in which this idiot can express feelings.
Last but not least, I’ve to thank Armindo, he welcomed me in his lab after few hours of discussions
about science. I had started a PhD with the objective of learning how to do science, and under his
mentorship I matured from the raw researcher I was. We had very interesting discussions and
exchanged so many crazy ideas along these 5 years and we managed to carry at the end this
xiii
Index
Declaration-Declaração ... vii
Financial Support
–
Apoio Financeiro ... ix
Acknowledegments ... xi
Index ... xiii
Abstract ... xvii
Resumo ... xix
List of Figures ... xxi
List of Tables ... xxiii
List of Abbreviations ... xxv
Chapter 1 | General Introduction ... 1
Oxidative stress: a changing paradigm ... 2
Reactive oxygen species: why hydrogen peroxide? ... 3
Hydrogen peroxide targets: Cysteines ... 4
Hydrogen peroxide concentrations
in vivo
... 6
Antioxidant defenses ... 9
Peroxiredoxins ... 9
Thioredoxin and Thioredoxin Reductase ... 13
Sulfiredoxins ... 15
Alternative defense mechanism ... 16
Hydrogen peroxide signal processing... 17
The Peroxiredoxin/Thioredoxin/Thioredoxin Reductase System as hydrogen peroxide sensor. ... 17
Aim of the thesis ... 20
Chapter 2 | Design principles for thiol redox signaling: mapping the phenotypic repertoire of the cytoplasmic 2-Cys peroxiredoxin – thioredoxin system ... 21
Abstract ... 22
Introduction ... 22
Model Formulation ... 25
Results ... 26
A phenotypic map of the PTTRS ... 26
Identifying the best region for signaling and protection ... 31
Responses to stress ... 33
xiv
Discussion... 38
Material and Methods ... 40
Chapter 3 | Hydrogen peroxide concentrations classifier ... 41
Abstract ... 42
Introduction ... 42
Results ... 43
Genetic Comparators Digitize Analog Gene Expression ... 43
Complex Signal Processing Circuits Composed of Genetic Comparators ... 47
A Mixed-Signal Processing Gene Circuit ... 52
Discussion... 53
Material and Methods ... 56
Strains and plasmids... 56
Circuit characterization ... 56
Chapter 4 | Measuring hydrogen peroxide concentrations in vivo ... 57
Abstract ... 58
Introduction ... 58
Results ... 59
Discussion... 60
Material and Methods ... 61
Strains and bacteria preparation ... 61
Zebrafish Husbandry... 61
Injections ... 62
Fin-fold amputation ... 62
Image acquisition ... 62
Imaging ... 62
Distance analysis ... 62
Chapter 5 | General discussion and future perspectives ... 63
Appendix A | Design principles for thiol redox signaling: mapping the phenotypic repertoire of the cytoplasmic 2-Cys peroxiredoxin – thioredoxin system supplementary information ... 65
The systems design space methodology for characterizing the phenotypic repertoire of
biochemical circuits ... 66
Design space analysis of the PTTRS model ... 74
Performance criteria ... 76
xv
PTTRS Topologies ... 85
Topology A-i ... 86
Topology A-ii ... 87
Topology A-iii ... 87
Topology A-iv ... 88
Topology B-i ... 88
Topology B-ii ... 89
Topology B-iii ... 89
Topology B-iv ... 90
Topology C-i ... 91
Topology C-ii ... 91
Topology C-iii ... 92
Topology C-iv ... 93
Dichotomy threshold Calculation ... 93
Parameters Estimations ... 94
Estimation of protein concentrations from proteomic datasets ... 94
Jurkat T Cells ... 94
Peroxiredoxins concentration and rate constants ... 94
Thioredoxin concentration and rate constant ... 96
Thioredoxin Reductase concentration and activity ... 96
H2O2 Permeability ... 97
Alternative H2O2 sinks ... 98
Sulfiredoxin concentration and activity ... 99
Other cells ... 101
PTTRS parameters other cell line ... 102
Design space PTTRS other cell lines ... 103
Appendix B | Hydrogen peroxide concentrations classifier supplementary information ... 105
Supplementary figures ... 105
Plasmids and synthetic parts ... 132
Data Processing and Calculations ... 191
Calculating the sigmoidal fit, input threshold, and relative input range ... 192
xvi
Calculating the relative resolution of a genetic analog-to-digital converter circuit ... 195
xvii
Abstract
Reactive oxygen species (ROS) such as hydrogen peroxide (H2O2), are now known to play critical roles in signal transduction and in coordinating key cellular processes. However, these species can
also covalently damage macromolecules and originate other even more deleterious compounds.
At the core of this twine between signaling and defense lays the Peroxiredoxin Thioredoxin
Thioredoxin Reductase (PTTR) system. Experimental studies of the PTTRS highlighted many
commonalities among different types of cells and organisms, but also intriguing differences in cells’ responses to hydrogen peroxide.
The current work aims to study the PTTR system and its characteristics. Using a minimal
mathematical model, we seek to uncover the general principles of how organisms exploit the
properties of ROS for regulation of other protein while avoiding their deleterious effects.
These principles, in the form of relationships among rate constants and species concentrations, are
thoroughly supported by experimental observations in a variety of organisms and allow to correlate
proteins abundance patterns with the modes of response.
Depending on the relative abundances of peroxiredoxins, sulfiredoxin, thioredoxin, thioredoxin
reductase and alternative H2O2-consuming proteins, the system is capable of distinct responses to changing hydrogen peroxide supplies, including proportional, ultrasensitive, and hysteretic (toggle
switch) ones.
The complete characterization of the system however requires the definitions of the operative
conditions in which the organism lives. A major and so far not univocally defined value is the
maximum attained hydrogen peroxide concentration in vivo. To address this problem were
developed a series of sensor with different thresholds and capable of memory functions. The
peroxide classifier was then used in an inflammation animal model to measure the maximum
attained concentrations.
The mathematical model developed in this system and the studies of the general principles
underlying the PTTR system together with the experimental application of the H2O2 classifier could be used in clinical research or drug development.
xix
Resumo
Várias espécies reactivas de oxigénio (ROS) , tal como o peróxido de hidrogénio (H2O2), foram recentemente identificados como modeladores de sinalização e coordenação de importantes
processos celulares. No entanto, estas partículas podem causar danos oxidativos em
determinadas macromoléculas ou até originar outros metabolitos ainda mais reactivos.
Central a todo este processo de equilíbrio entre sinalização e dano oxidativo, encontra-se o
importante sistema de defesa redox Peroxiredoxina Tioredoxina Tioredoxina-Reductase
(PTTR). Várias evidências experimentais envolvendo o sistema PTTR, apontam para um grande
nível de conservação entre diferentes tipos de células e organismos, mas no entanto é também
evidente alguma disparidade em termos de resposta ao stress induzido por H2O2.
Este trabalho foi desenvolvido visando estudar o sistema PTTR. Através de um modelo matemático
minimalista, procurámos descobrir os princípios base de como os organismos utilizam os ROS
na regulação de outras proteínas de maneira a evitar os seus efeitos nefastos.
Os pricípios base foram desenvolvidos sob a forma de relações entre as constantes de reacção e
a concentração das espécies. Princípios esses que foram solidamente apoiados por observações
experimentais em diferentes organismos, tornando possível uma correlação clara entre o padrão
de expressão das proteínas e a forma como o organismo responde ao stress.
Dependendo das concentrações relativas de peroxiredoxinas, sulfiredoxina, tioredoxina,
tioredoxina-reductase e outras proteínas que alternativamente consomem o H2O2, o sistema é capaz de responder de forma diferente a mudanças de H2O2, incluíndo alterações proporcionais, ultra-sensíveis e histeresicas.
No entanto, a caracterização do sistema requer o conhecimento das condições oxidativas nas
quais o organismo se desenvolve. Para isso, desenvolvemos uma série de sensores de H2O2 capazes de detectar diferentes níveis de exposição e capazes de memorizar esse mesmo
contacto. O sensor foi posteriormente validado num modelo animal de inflamação de maneira a
determinar os níveis máximos de exposição in vivo.
Nest trabalho desenvolveu-se um modelo matemático que em combinação com o sensor, podem
xxi
List of Figures
Figure 1.1| Different sources of hydrogen peroxide in eukaryotic cells. ... 4
Figure 1.2| Cysteine biochemistry allows for redox-dependent signaling. ... 6
Figure 1.3| Schematic classification of the oxidative stress based on intensity. ... 7
Figure 1.4| Hyper ratio in COS-7 cells exposed to different concentration of H2O2. ... 8
Figure 1.5| Phylogenetic tree of the peroxiredoxin family. ... 9
Figure 1.6| 2-Cys Peroxiredoxins catalytic cycle. ... 11
Figure 1.7| Typical 2-Cys peroxiredoxin quaternary structures. ... 12
Figure 1.8| Redox reactions catalyzed by a mammalian Trx system comprising thioredoxin
reductase (TrxR), thioredoxin (Trx) and NADPH. ... 13
Figure 1.9| Phylogenetic relationships between high molecular weight thioredoxin reductase
(H-TrxR), low molecular weight Thioredoxin reductase (L-TrxR). ... 13
Figure 1.10| Phylogeny of Thioredoxin homologs from representative species of the three domains
of life. ... 14
Figure 1.11| Relatedness tree for Sulfiredoxin sequences. ... 15
Figure 1.12| Comparison of the proposed sulfinic acid reduction mechanism of Sulfiredoxin. ... 16
Figure 1.13| Trx-Ask1 interaction. ... 18
Figure 1.14| Model for the production, signaling role, and removal of H2O2 in growth factor-stimulated cells. ... 19
Figure 2.1| The peroxiredoxin / thioredoxin / thioredoxin reductase system (PTTRS) model. ... 23
Figure 2.2| Allowed topologies of the various regions of distinct behavior of the PTTRS in the
parameters space for biologically plausible conditions. ... 34
Figure 2.3| Design space of the PTTRS for human Jurkat T cells and erythrocytes. ... 36
Figure 2.4| Gene regulation of the PTTRS components to induce adaptation. ... 38
Figure 3.1| Comparator overview. ... 43
Figure 3.2| Genetic comparators with different activation thresholds. ... 46
Figure 3.3| Bandpass filters assembled from low-pass and high-pass filters. ... 48
Figure 3.4| Multi-bit analog to digital converters. ... 51
Figure 3.5| Mixed-signal computation and concentration-dependent logic. ... 53
Figure 4.1| Measuring hydrogen peroxide gradients in zebrafish. ... 59
Figure 4.2| Sensors activation between two wounds ... 60
Figure A.1| PTTRS model. ... 66
Figure A.2| Dimensionless PTTRS model. ... 69
Figure A.3| Fit to the time course of NADPH consumption reported in Figure 6A of ref. [24]. ... 96
xxii
Figure A.5| Sulfiredoxin activity estimation from fit of ref. [121] ... 100
Figure A.6| Design space of the PTTRS for other cell lines. ... 103
Figure B.1| Analog H2O2–sensor ... 105 Figure B.2| Digitization of an analog input by inverting target DNA on a medium-copy plasmid (MCP)
versus a bacterial artificial chromosome (BAC). ... 107
Figure B.3| Feedforward cascade involving a recombinase-invertible trans-acting transcriptional
element on a BAC. ... 108
Figure B.4| Amplifying BAC output with Copy Control. ... 110
Figure B.5| Flow cytometry histograms for comparators with different activation thresholds. ... 112
Figure B.6| A bandpass filter assembled from a low-threshold high-pass circuit and a
medium-threshold low-pass circuit. ... 116
Figure B.7| A bandpass filter assembled from a low-threshold high-pass circuit and a high-threshold
low-pass circuit. ... 119
Figure B.8| Ternary Logic. ... 122
Figure B.9| 2-bit Analog-to-digital Converter. ... 123
Figure B.10| Mixed-signal processing and concentration-dependent logic. ... 124
Figure B.11| Digital-to-analog converters and analog-to-digital converters are complementary
systems that translate digital signals to analog signals, and vice versa. ... 125
Figure B.12| Growth curves for cells containing 0, 1, 2, or 3 recombinases at different concentrations
of H2O2. ... 127
Figure B.13| Growth curves for cells containing 0, 1, or 2 recombinases on 2 plasmids or 2
recombinases on 1 plasmid at different concentrations of H2O2. ... 129
Figure B.14| Scale-up of the 2-bit ADC circuit. ... 130
xxiii
List of Tables
Table 1.1| Some known sources of oxidative stress. ... 2
Table 1.2| Acidity and Hydrogen Peroxide reactivity of several protein thiolates. ... 5
Table 1.3| Summary of Prx Subfamily phylogenetic distribution and structures. ... 10
Table 1.4| kinetic parameters for Thioredoxin from different organisms ... 15
Table 2.1| Biologically relevant regimes represented by the dominant species in that region and the
consequent scheme of the system. ... 27
Table 2.2| Evaluation of the local performance in all biologically relevant Regions. ... 32
Table 2.3| Topology Superfamilies definitions. ... 35
Table 4.1| Sensor activation thresholds. ... 59
Table A.1: Dimensionless Groups ... 67
Table A.2| Phenotypic regions inequalities. ... 74
Table A.3| Biologically plausible phenotypic region steady state. ... 76
Table A.4| Performance criteria evaluated for the biological plausible regions ... 77
Table A.5| Minimized physiologically plausible regimes inequalities. ... 85
Table A.6| Cell lines kinetic parameters. ... 102
Table B.1| List of plasmids used in experiments ... 134
Table B.2| Plasmid Sequences. ... 135
Table B.3| List of synthetic parts ... 189
xxv
List of Abbreviations
List of abbreviations in alphabetical order.
Abbreviation Complete name
ADC Analog to digital converter
AhpC Alkyl hydroperoxide reductase
Akt Protein kinase B
ASK-1 apoptosis signaling kinase-1
aTc anhydrotetracycline
ATP Adenosine triphosphate
BAC Bacterial artificial chromosome
BFP Blue fluorescent protein
BOS Basal oxidative stress
Cat Catalase
CC Copy control
Cys Cysteine
DAC Digital to analog converter
E.coli Escherichia coli
EGF Epidermal growth factor
FF Fully folded
GFP Green fluorescent protein
GMA Generalized Mass Action
GPx Glutathione peroxidase
GR Glutathione Reductase
Grx Glutaredoxin
GSH Glutathione
H2DCF 2′,7′-dichlorodihydrofluorescein
H2O2 Hydrogen peroxide
HCP High-copy plasmid
HO• Hydroxyl radical
HOS High oxidative stress
hpf Hours post fertilization
hPrxI Human Peroxiredoxin I
hPrxII Human Peroxiredoxin II
xxvi
hpw Hours post wounding
IL Interleukin
IOS Intermediate oxidative stress
LB medium Luria-Bertani medium
LCP Low-copy plasmid
LOS Low oxidative stress
LU Locally unfolded
MAPK/ERK mitogen-activated protein kinases, originally called extracellular signal-regulated kinases
MCP Medium-copy plasmid
Met Methionine
NADPH Nicotinamide adenine dinucleotide phosphate
NF-κB Nuclear factor kappa-light-chain-enhancer of activated B
cells
NOX NADPH-oxidase
oO2•
- Superoxide anion
ODE Ordinary differential equations
Orp1 Oxidant receptor protein
OS Oxidative stress
PDGF Platelet-derived growth factor
PI3K phosphatidylinositol 3-kinase
PIP PI 3,4,5-trisphosphate
PPP Pentose Phosphate Pathway
Prx Peroxiredoxin
PTEN Phosphatase and tensin homolog
PTP-1B Protein-tyrosine phosphatase 1B
PTTRS Peroxiredoxin-Thioredoxin-Thioredoxin Reductase System
RBS Ribosome binding site
RFP Red fluorescent protein
ROS Reactive oxygen species
S. pombe Schizosaccharomyces pombe
SOD Superoxide dismutase
Srx Sulfiredoxin
STAT3 Signal transducer and activator of transcription 3
TNFα Tumor necrosis factor α
xxvii
TrxR Thioredoxin reductase
TSAI Thiol-specific antioxidant protein 1
TSAII Thiol-specific antioxidant protein 2
2
Oxidative stress: a changing paradigm
The definition of oxidative stress (OS) has been changing over the years, due to a shift in the
paradigm of the processes that involve some reactive oxygen species (ROS): from deleterious to
necessary in the normal functions of the organism.
A comprehensive definition of this phenomenon (given by H. Sies and D. Jones) reported OS as:
“an imbalance between oxidants and anti-oxidants in favor of the oxidants, leading to a
disruption of redox signaling and control and/or molecular damage” [1,2]. This definition highlights the importance of sensing oxidation sources, transmitting information carried by them,
and activating proper responses. Such responses may eventually prevent the propagation of
damage to key cellular components such as proteins, nucleic acids and lipidic membranes.
The OS sources can be endogenous (ensuing from mitochondrial respiration, protein folding
autoxidation, etc.) or external to the organism. The latter may be originated by different factors
(Table 1.1).
Table 1.1| Some known sources of oxidative stress.
Condition Proposed Source Likely reactive species
produced
Hyperoxia;
hypoxia;
ischemia;
reperfusion
Mitochondria; NADPH oxidases;
xanthine oxidase; nitric oxide
synthases
Superoxide,
hydrogen peroxide,
nitric oxide,
peroxynitrite
Inflammation Phagocyte NADPH oxidase;
myeloperoxidase; inducible
nitric oxide synthase
HOCl, chloramines,
HOBr, bromamines,
HOSCN, oxyradicals,
nitrogen dioxide,
carbonate radical,
nitric oxide, peroxynitrite
Activation of receptors to
agonists (e.g. Fas, TNF-α,
angiotensin II)
NADPH oxidases; mitochondria;
nitric oxide synthases
Superoxide,
hydrogen peroxide,
nitric oxide
Xenobiotic metabolism Peroxidases; flavoprotein
reductases; autoxidation
Oxyradicals,
superoxide,
hydrogen peroxide
Table 1.1 from ref. [3]
Deregulation of redox signaling pathways, with consequent OS, has been shown to be involved into
the development of pathologies such as cardiovascular disease[4], inflammatory bowel disease
[5,6], atherosclerosis, diabetes and metabolic disease [7,8] and neurodegenerative disease (e.g.
3
OS play also a major role in tumor incidence and progression, where the antioxidant defenses are
necessary for the initiation and the survival of the cancer [10,11]. Furthermore, in higher organisms
the aging phenomenon is associated with an increase in the basal level of oxidants, called
inflammaging [12].
It is thus important to analyze the signaling/defense system to better understand how to exploit the
underlying mechanism of redox regulation for developing new drugs that target the diseases listed
above.
Reactive oxygen species: why hydrogen peroxide?
ROS are a heterogeneous family containing both highly reactive oxygen radicals (e.g., (O2) hydroxyl radical (HO•)) and less reactive non-radical oxidants (e.g., hydrogen peroxide H2O2, singlet oxygen (1O
2)).
In the former group we find radicals like HO•, which is a very strong and indiscriminate oxidant. It can react with many amino acid side chains with diffusion-limited rate constants. The short lived
nature of this ROS (estimations report 9
10 s at 37ºC [13]) spatially limit its interactions to the region where is generated.
HO• radicals are generated by the reaction of transition metal ions (e.g. ferrous, cuprous) with H2O2
or via the iron catalyzed reaction between O2 and H2O2.
•- •
-2 2 2 2
O H O HO OH O (1.1)
2+ - 2+
2 2
H O +Fe
HO +HO
Fe
(1.2)This can induce extensive damage especially in cells carrying heme groups[14].
O2 is in general moderately reactive with biological macromolecules, but it can generates very
reactive radicals such as HO and peroxinitrite [15]. Additionally, it is very reactive with iron-sulfur
clusters of dehydratases, inactivating these enzymes and releasing Fe2+ in the process [16,17].
O2 also reacts with reduced glutathione leading to the formation of sulfinyl and thiyl radicals to then regenerating itself in a self-sustaining cycle[18]. Furthermore, it promotes DNA damage [17]
O2 is the primary product of NADPH oxidases [19] (NOX) and a byproduct of the mitochondrial respiration [20]. Because of its charged nature at physiological pH it can only permeate cell
membranes through anion-channels [21]. O2 is removed mainly by dismutation to O2 and H2O2.
in a reaction catalyzed by superoxide dismutase (SOD), with a catalytic rate constant of
approximately 109 M-1s-1 [22,23].
Nearly all oxygen tolerant organisms contain SOD isoforms [24] that tightly control the intracellular
concentration of O2.In addiction to this, the O2 limited membrane permeability describe a molecule with considerable limitation as signaling compound.
On the other hand, H2O2 possess crucial characteristics to behave as a redox messenger.
4
(e.g. PDGF, p53, thyrotropin, EGF, insulin, TNF-α) [19,25–29], presence of chemicals and immune (e.g. wound, inflammation) [30,31] or competitive responses (e.g. lactic-acid bacteria suppress
competition by excreting H2O2)[32].
H2O2 is a small uncharged compound that can diffuse through the cell membrane either via passive diffusion or aquaporin channels [33], due to the high similarity with the water dipole. As a matter of
fact organism have developed strategies to control this passive diffusion and to regulate the
composition of the membrane[34] to make it harder to cross in response to sustained oxidative
insults.
H2O2 and O2•- have a major role in redox biology, but considering the rapid conversion of the latter in H2O2 and the characteristic that this last has. We will focus on H2O2 as the principal ROS member, for signaling purposes.
Figure 1.1| Different sources of hydrogen peroxide in eukaryotic cells. Hydrogen peroxide can be
produced extracellularly, for example by the immunoglobulin G-catalyzed oxidation of water, by receptor/ligand interactions, and by phagocytic immune cells. O2•- , is produced by the partial reduction of the oxygen by
cytochrome c oxidase in the mitochondria, by membrane associated NADPH oxidase, or by 5’-lipoxygenase in the cytoplasm, it is then rapidly converted to H2O2 by the action of cytoplasmic and mitochondrial SOD.
Growth factor, cytokines and integrins stimulate the activation of NADPH oxidase and/or 5’-lipoxygenase. Figure and legend adapted from ref [35]
Hydrogen peroxide targets: Cysteines
Cysteines (Cys) are the most nucleophilic amino acids in proteins, showing (together with
methionine) a predisposition to oxidative modifications [36,37]. As a matter of fact, different
enzymes families use these amino acids transformations as regulation (e.g. proteases,
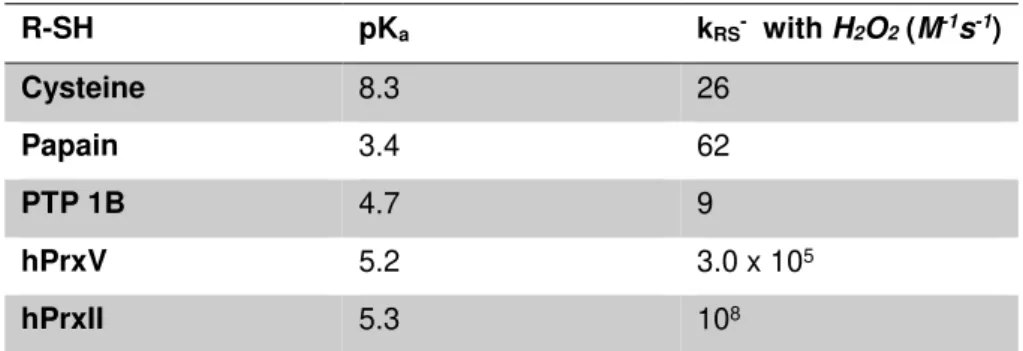
peroxidases, oxidoreductase [38]). The reactivity of a Cys thiol group, with H2O2 is dependent on the surrounding microenvironment and its acidity (pKa). A free Cys in the cytoplasm has a pKa between 8 and 9, which leaves the thiol group protonated and mostly non-reactive at physiological
5
the acidity to result in a pKa as low as 4 to 5, deprotonating the thiols (R-S-)[40]. However, the enhanced reactivity is not only correlated with the deprotonation but also depends on the
stabilization of the thiolate , that increase its nucleophilicity [40].
Table 1.2| Acidity and Hydrogen Peroxide reactivity of several protein thiolates.
R-SH pKa kRS- with H2O2(M-1s-1)
Cysteine 8.3 26
Papain 3.4 62
PTP 1B 4.7 9
hPrxV 5.2 3.0 x 105
hPrxII 5.3 108
Table 1.2 adapted from ref [40]
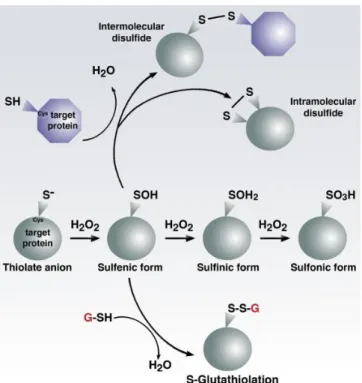
Cys have several different post translational modification (Figure 1.2) depending on the oxygen
degree of oxidation. The thiolate anion (R-S-), the reactive form of Cys, may upon interaction with
H2O2 generate the following oxidative modifications: sulfenic acid (R-SO-), sulfinic acid (R-SO2-),
sulfonic acid (R-SO3- ).
-
-2 2 2
H O +R-S
R-SO +H O
(1.3)2 2 2 2
H O +R-SO
R-SO +H O
(1.4)2 2 2 3 2
H O +R-SO
R-SO +H O
(1.5)Protein Cys sulfenates can condense with protein thiols or low-molecular-weight thiols such as
glutathione (glutathionylation) forming disulfides (henceforth denoted by R-SS-R or R-SSG,
respectively):
-
-2
R-S
H
R -SO
R-S-S-R +H O
(1.6)
-2
GSH R-SO
R-SSG+H O
(1.7)Thiol-disulfide oxidoreductase such as thioredoxin [41] or glutaredoxin [42] are able to resolve
exposed disulfide bonds at the expense of reducing equivalents. However, it is possible that some
cross-links are sterically hindered and thus irreversible.
Glutathionylation modifications are only resolvable through the glutaredoxins pathway [42,43].
Although R-SO2- of typical 2-Cys peroxiredoxins can be reduced at the expense of ATP and
6
Figure 1.2| Cysteine biochemistry allows for redox-dependent signaling. Specific reactive cysteine (Cys)
residues within target proteins can be modified by oxidative stress. Thiolate form can be progressively oxidized by reacting with H2O2 in: sulfenic, sulfinic, sulfonic form. The sulfenic form (SO-) is highly reactive and can
condensate with another thiol forming an intra- or inter molecular disulfide or with a glutathione molecule. Higher states of oxidation generally, but not always lead to irreversible modifications. Figure from ref. [47]
These reactions that lead to irreversible or non-repairable modifications are thus of particular
concern because they may generate protein aggregates or misfolding leading to toxic effect for the
organism. It is thus fundamental for an organism to maintain a tight control on the peroxide
concentrations. As a matter of fact H2O2 has been reported to be a possible mutagenic source by damaging DNA already with sub-micromolar intracellular concentrations [48].
Hydrogen peroxide concentrations
in vivo
Although high levels of H2O2 and other ROS generate cellular damage, it is becoming clear that low levels of H2O2 participate in cellular signaling to maintain homeostasis[49]. Despite the importance of H2O2 to cellular activities, the molecular mechanisms of its production, accumulation, function, and scavenging remain insufficiently understood. This is due to a large extent to the lack of
knowledge of the actual concentrations and fluxes of H2O2 in vivo, and to persistent uncertainties about the roles of distinct antioxidant defenses in physiological context.
A wide range of peroxide concentration has been used in experiments to study the system (from
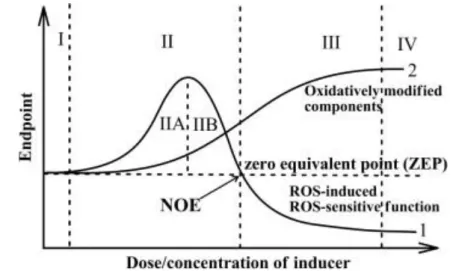
µM to mM) with different and sometime subjective classifications. Lushchak [50] tried to give the
following formal semi-quantitative definition of OS based on the observable phenotype produced
(Figure 1.3).
Under basal oxidative stress (BOS) there are no observable outcomes. This state represents the
normal functioning of the cell. An increase in the oxidative load will shift the organism to low-intensity
oxidative stress (LOS), characterized by oxidation of the most reactive cellular components and
induction of the redox-dependent response. Intermediate-intensity oxidative stress (IOS) is high
7
inactivation of antioxidant and associated enzymes that were upregulated in the LOS). This will
generate an apparent negative response of the ROS- induced functions to increasing
concentrations of the oxidant and an even higher oxidation of the available targets. Finally, in the
high-intensity oxidative stress (HOS)virtually all available potential substrates are oxidized [50].
Figure 1.3| Schematic classification of the oxidative stress based on intensity. This figure shows the
behavior of the redox couples (curve 2) and of the ROS dependent response (curve 1) across different intensity of OS. I – basal oxidative stress zone (BOS); II – low intensity oxidative stress (LOS); III – intermediate intensity oxidative stress (IOS); and IV – high intensity oxidative stress (HOS). Figure from ref. [50]
A sensitive and precise determination of H2O2 levels in vivo is essential to examining the role of H2O2 in physiological or pathological processes[51]. The acquisition of such knowledge has been delayed due to the difficulty of quantifying and tracking the small, diffusible and fast cleared H2O2 molecules in living cells. Various methods are nowadays available for the measurement of the H2O2 concentration.
A category of methods sensitive enough to determine physiological H2O2 concentrations is based
on dihydro compounds such as 2′,7′-dichlorodihydrofluorescein (H2DCF) that fluoresce upon oxidation. They are widely used because of their sensitivity and simplicity, but these probes lack
specificity, reacting with a variety of ROS including nitric oxide, peroxynitrite, and hypochloride in
addition to H2O2 [52].
Another example are deprotection reaction-based probes that fluoresce upon H2O2-specific removal of a boronate group, rather than on nonspecific oxidation [53–55]. Intracellular H2O2 production can be also visualized by highly H2O2-specific, genetically encoded, and reversible fluorescent constructs, whose characteristics enable in vivo real-time dynamic H2O2 determinations.
Two main genetically encoded sensors are currently available. One is HyPer [56,57] and the other
is Orp1-redox-sensitive green fluorescent protein 2 (roGFP2) [51]. In the latter, the reaction between Orp1 peroxidase and H2O2 results in a disulfide that changes the roGFP2 β-barrel structure, leading to changes in the spectrum of the fluorescent protein. Analogously, Hyper
consists of a circularly permuted yellow fluorescent protein inserted into the regulatory domain of
8
[59]. The disulfide formation leads to conformational changes and hence to a change in the YFP
excitation spectrum (400nm/500nm excitation and 516nm emission). The readout in both cases
(roGFP2 and Hyper) is the ratio between the emission of light by the protein when excited with one
or the other wavelength. In vitro assays for Hyper reported a ratio between 1.5-3.3 respectively to
25-250 nM of H2O2 [56]. A calibration curve, performed by FACS, over Hyper-expressing COS-7 cells exposed to various H2O2 concentrations showed a minimum external concentration of 5 µM to activate the sensor and a saturation at around 20 µM
Figure 1.4| Hyper ratio in COS-7 cells exposed to different concentration of H2O2. The ratio between the
fluorescence excited by 488 nm and 405 nm lasers as a function of H2O2 concentration. In red are reported
the experimental points in black the fitting curve ( ; 4.5, 3.7, 9.7, 2.8 1
A D
y D A B C D
x C
). Figure and
legend adapted from ref. [56].
The development of these intracellular redox sensors evolved over the past years improving their
performances [57] and eventually being used in animal model. The study of the inflammation-like
response in zebrafish performed by Niethammer et al. [61] showed the importance of H2O2 in the wound healing process and the establishment of a gradient in the tissue (due to NADPH-oxidases)
that would function as chemotaxis signal for the immune system cells[30,57,62]. Both these work
from Niethammer et al. and Pase et al. (respectively ref. [61] and ref. [30]) estimated, on the calibration curve previously done (ref. [56] and Figure 1.4), a concentration that ranged from 5-50
µM having the highest value of the gradient at the wounding site. This measurement unfortunately
lacks a proper calibration. The cells examined are from various tissue and heterogeneous in
antioxidant defense expressions and redox state. Furthermore, the calibration curve was calculated
on human COS-7 cell. It spans over the entire linear region of the sensor possibly entering the
saturation region. The Hyper mRNA injected can be degraded at different rates in different cells
giving varying Hyper expressions.
Other estimations made on the data available in literature calculate the H2O2 concentration in the human plasma being between 1-5 µM in normal condition and increasing up to 30-50 µM in chronic
9
The lack of consensus about the H2O2 concentration, together with the wide dynamic range covered by various experimental set-up (from µM-mM of external H2O2) make of the physiological oxidative load a puzzle that has still to be properly addressed.
Antioxidant defenses
Organisms have developed a variety of antioxidant defenses to counterbalance and control the
ROS compounds. There are two major categories of defense: enzymatic (e.g. peroxidase, catalase,
superoxide dismutase etc.) and non-enzymatic (e.g. vitamin C, A) [65]. In the following paragraphs
we will focus on the Peroxiredoxin-Thioredoxin-Thioredoxin Reductase system (PTTRS), which is
one of the main subjects of interest in this thesis
Peroxiredoxins
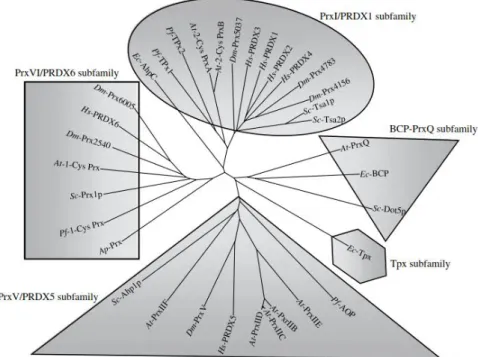
Peroxiredoxins (Prx) are a Cys-based class of scavengers for H2O2, which protect the cells from oxidative insults and prevent damage to cellular key components. Discovered to be ubiquitously
present in several organisms, from archaea to humans [66], these proteins show abundances,
structures similarities and properties that are conserved even amongst kingdoms [67] (Figure 1.5).
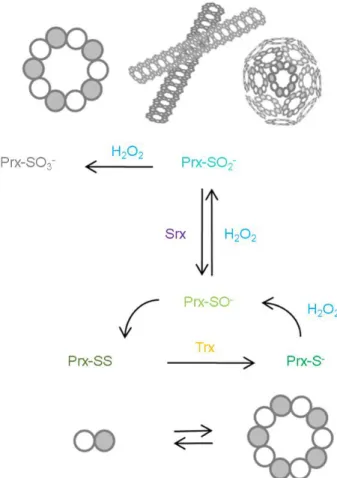
Their catalytic cycle involves the oxidation of a peroxidatic Cys thiolate (CP-S-), located in a universally conserved PXXXTXXC motif, to sulfenic acid (CP-SO-). This sulfenic acid eventually reacts with a resolving cysteine (CR) forming an inter- or intra- molecular disulfide that will be reduced restoring the thiolate.
Figure 1.5| Phylogenetic tree of the peroxiredoxin family. Protein alignment was performed with clustalX
Dm-10
Prx5037 (NM_079663); Dm-PrxV (NM_176513); Dm-Prx6005 (NM_078739); Dm-Prx2540 (NM_165769); PRDX1 (NM_002574); PRDX2 (NM_005809); PRDX3 (NM_006793); PRDX4 (NM_006406); Hs-PRDX5 (NM_012094); Hs-PRDX6 (NM_004905). Figure and legend from ref [68].
Prx are divided into six different families depending on the mechanism of resolution of the sulfenic
acid and their oligomeric state.
Table 1.3| Summary of Prx Subfamily phylogenetic distribution and structures.
Subfamily Phylogenetic
distribution
Structural distinctions
relative to Prx core fold
Oligomeric states and
interfaces
Prx1/AhpC
Archaea, bacteria,
plants and other
eukaryotes
Extended C terminus
B-type dimers, (α2)5
decameres (and rare (α2)6
dodecamers) through A
interface
Prx6
Archaea, bacteria,
plants, and other
eukaryotes
Long, extended C
terminus
B-type dimers, some (α2)5 decamers through A interface
AhpE Bacteria Extended loop at N
terminus A-type dimers
PrxQ Archaea, bacteria,
plants and fungi Extended helix α5
Monomers and A-type dimers
Tpx Bacteria N-terminal hairpin A-type dimers
Prx5
Bacteria, plants,
and other
eukaryotes
Pi helix insertion in β;
~20% fused with Grx
domain
A-type dimers
Table 1.3 adapted from ref [69]
Prx1/AhpC and Prx6 subfamilies have the widest biological distribution ( Table 1.3, Figure 1.5). The
former are very abundant in cells, as illustrated by the following examples. They compose
~0.1%-1% of the soluble protein in rat and human cells [70], with peroxiredoxin II (hPrxII) being the third
most abundant protein in human erythrocytes [71]. Thioredoxin peroxidase I (TSA1) constitutes the
~0.2-0.7% of the total soluble protein in S. cerevisiae [72]. Alkyl hydroperoxide reductase (AhpC) accounts for 0.4% of the proteome of Escherichia coli according to data from [73,74], and it is annotated amongst the ten most expressed proteins in this organism [75–77]. In eukaryotic cells, Prx1/AhpC peroxiredoxins are mainly located in the nucleus and cytoplasm.
The conserved fold at the active site of these proteins grant them high catalytic rates for the
reduction of H2O2: hPrxII shows a second order rate constant of 108 M-1s-1 [78] , TSA1 of 2.2x107 M-1s-1 [79], AhpC of 4x107 M-1s-1 [80].
The high abundances together with the high catalytic rates of ~106-108 M-1s-1 for H2O2 reduction may account, in the absence of inhibiting factors[81], for the consumption of more than the 90% of
11
Figure 1.6| 2-Cys Peroxiredoxins catalytic cycle. H2O2 oxidizes the peroxidatic cysteine to a sulfenic acid
(Prx-SO-), which then condenses with the resolving cysteine from an adjacent monomer to form a disulfide (PrxSS). This step requires first a local unfolding at the active sites that rearranges the peroxidatic and resolving cysteines regions. The cycle is then closed by the reduction of PrxSS often carried out by thioredoxin (Trx), at the expenditures of a reducing equivalent (NADPH) supplied through thioredoxin reductase (TrxR) eventually returning Prx to its fully folded structure.
Peroxiredoxins of the Prx1/AhpC subfamily, commonly referred as typical 2-Cys peroxiredoxins,
are pentamers of dimers. The subunits in each dimer are arranged in an antiparallel fashion with
the CP of one dimer facing the CR of the other. These peroxiredoxins reduce H2O2 through a three-step cycle (Figure 1.6) that requires the formation of an inter-subunit disulfide bond, and that is
maintained by the supply of reducing equivalents through the thioredoxin system.
Eukaryotic 2-Cys peroxiredoxins of the Prx1/AhpC subfamily, such as human peroxiredoxins I
(hPrxI) and II (hPrxII), are susceptible to inactivation by their own substrates due to the conversion
of their peroxidatic cysteines to sulfinic (Prx-SO2-) and sulfonic (Prx-SO3-) acids. This phenomenon, called hyperoxidation, is facilitated by two phylogenetically conserved structural motifs. Namely, a
GGLG and a C-terminal extension with a YF, perturbing the unfolding-folding equilibrium of the
active sites and thereby making them more prone to hyperoxidation[82]. The change in oxidative
state also translates into a rearrangement of the quaternary structure of the Prx, this is usually
associated with a change in function from scavenger to holdase/chaperone[83–85]. Prokaryotic peroxiredoxins are typically more resistant to hyperoxidation, with few exceptions[84,86]. They
require H2O2 concentrations in the mM range to be inactivated [82] while eukaryotic ones, as hPrxII,
are completely inactivated with 40 µM of H2O2 under similar conditions[87]. Prx-SO2- can be slowly
12
Figure 1.7| Typical 2-Cys peroxiredoxin quaternary structures. During the reduction process, the Prx
molecules alternate between dimeric and decameric states. The reduced, decameric form of the protein is the most reactive with H2O2. As the level of H2O2 increases, eukaryotic Prxs can react with a second H2O2
molecule to form the sulfinic acid form (Prx-SO2- ) and, as a result, are inactivated. This hyperoxidation stabilizes
the decameric state of the Prx molecule and can lead to the formation of filamentous and spherical, high molecular weight species, these play chaperon or holdase roles in the organism. Figure and legend adapted from ref [89]
Srx is a very inefficient enzyme[45,90,91], it is widespread among eukaryotes, but exceptions exist
where is not yet clear which enzymes recovers the sulfynilated form [92]. In several organisms
hyperoxidation occurs not only upon extreme insults but also under physiological conditions[93].
The wide phylogenetic conservation of the above-mentioned structural motives suggests that
sulfinylation confers properties that are favored by natural selection
Another possible evolutionary driving force that lead to this inactivation mechanism, is the so called
“floodgate hypothesis”. It proposes that a local inactivation of Prx allows H2O2 to increase locally thus propagating the signal to other target proteins that otherwise would be easily outcompeted by
Prx.[82]
Moreover, Day et al.[94,95] showed that in S. pombe the survival to mM concentration of H2O2 was diminished when Prx was not inactivated anymore, demonstrating the importance of this form.
The Prx6 family, comprehend the 1-Cys peroxiredoxins (e.g. Human PrxVI). they are mostly
cytosolic located, and their catalytic cycle requires GSH to be competed. In particular upon reaction
with H2O2 the active site thiolate is oxidized to a sulfonate, whose reduction is dependent on
13
role in H2O2 metabolism since from proteomics data we found its concentrations to be similar to those of hPrxI and hPrxII.
Thioredoxin and Thioredoxin Reductase
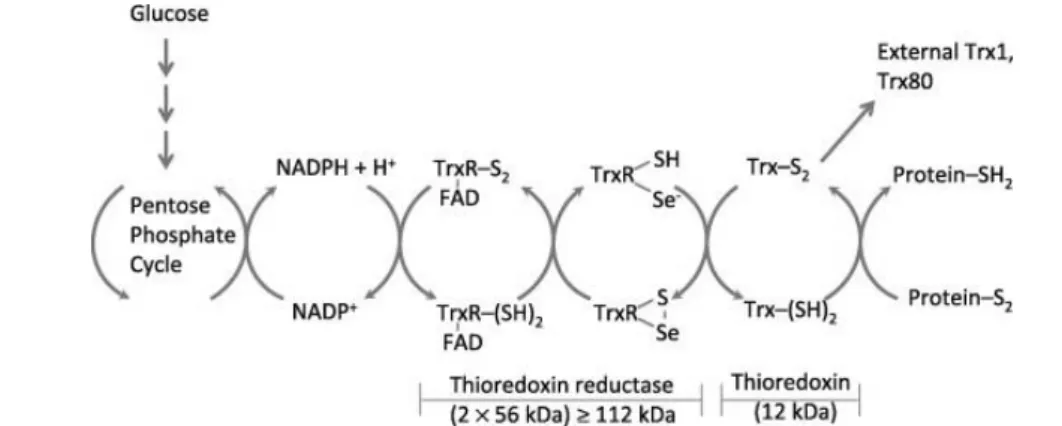
Thioredoxin (Trx), is a major disulfide reductase enzyme, widely distributed in all living organisms
from bacteria to mammals (Figure 1.10)[98]. Trx is characterized by a conserved active domain
Cys-Gly-Pro-Cys, which can reduce exposed disulfide substrate generating oxidized Trx [41]. The
reducing equivalents necessary to support this reaction are provided by the FAD-containing
enzyme thioredoxin reductase (TrxR).
Figure 1.8| Redox reactions catalyzed by a mammalian Trx system comprising thioredoxin reductase
(TrxR), thioredoxin (Trx) and NADPH. The electron source of the Trx system is NADPH, which is largely
produced from the pentose phosphate pathway. The oxidized thioredoxin (Trx-SS) is reduced by NADPH and the selenoenzyme TrxR. Electrons are transferred from NADPH to FAD, then to the N-terminal redox active disulfide in one subunit of TrxR, and finally to the C-terminal active site Gly–Cys–Sec–Gly of the other subunit[99]. Reduced thioredoxin (Trx-(SH)2) catalyzes disulfide bond reduction in many proteins. Figure and
legend from ref. [100]
The reduction of Trx is carried out by a selenoenzyme TrxR. Two types TrxRs have been
characterized, both belong to the flavoprotein family and both function as homodimers. The
monomers possess a FAD prosthetic group, a NADPH-binding site and an active site [101].
However, the two groups are different in amino acid sequences and catalytic mechanisms
[101,102], showing only a ~20% sequence identity [103].
Figure 1.9| Phylogenetic relationships between high molecular weight thioredoxin reductase (H-TrxR),
low molecular weight Thioredoxin reductase (L-TrxR). In the figure the phylogenetic tree is also
14
mercuric reductase (MerR), lipoamidedehydrogenase (LipD), alkylhydroperoxide reductase F52A (AhpF). There is a large sequence divergence between the two TrxRs group and a complex gene history for all six enzymes. The tree was derived using maximum likelihood methods: 180 aminoacids aligned between different enzymes and used in phylogenetic inference. Scale bar represents inferred number of changes per site. Figure and legend from ref. [103].
The first TrxR type, is characterized by a high molecular weight (~55 kDa, H-TrxR), can be found
mostly in higher organisms such as Homo sapiens, C. elegans and Drosophila melanogaster but also in the malaria parasite P. falciparum [104–107]. Mammalian TrxRs consist of two dimers arranged in an antiparallel fashion [99], they use one reducing equivalent from NADPH per
molecule of Trx in a ping-pong type of reaction [102]. Bacteria, archaea, fungi and plants commonly
possess another TrxR type with a low molecular weight (~35 kDa, L-TrxR) [108].
The thioredoxin system (Trx, TrxR and NADPH) can provide electrons to a large range of enzymes
and was originally found to play a critical role in DNA repair and replication by being the reducing
substrate of ribonucleotide reductase (RNR), together with Grx. [109].
Figure 1.10| Phylogeny of Thioredoxin homologs from representative species of the three domains of
life. Branch lengths were estimated using maximum likelihood with rete variation modeled according to a
gamma distribution. Scale bar represents amino acid replacements per site per unit evolutionary time. Posterior probabilities are shown at nodes of the phylogeny when greater than 50%. The lack of strong node supports deep in the phylogeny results from the ambiguous placement of mitochondrial sequences, possibly due to long branch attraction effects with nonbacterial sequences. Figure and legend from ref. [98]
Eukarya possess two homologue thioredoxin systems, one cytoplasmatic and the other
mitochondrial [98]. In addition to the catalytic active site thiols, mammalian cytoplasmic Trx possess
three conserved thiols[110]. Two of these are closely located and can form an intramolecular
disulfide; the remaining is located at the surface of the protein and can be either
glutathionylated[111] or S-nitrosyilated[112] upon oxidative insults. The disulfide cannot be reduced
by TrxR and delays the reduction of the Trx main active site by the same enzyme[110]. The
presence of these other thiols groups has been postulated to be a further layer of control on Trx
activity. Perer-Jimenez et al. [98] analysed the kinetics of 8 phylogenetically different Trxs. They share a common Michaelis-Menten mechanism in which the substrate disulfide first reorient and
15
constant for this mechanism are of the same order of magnitude ~105 M-1s-1 (Table 1.4) and similar even between phylogenetic distant Trx like the human or the E. coli one (Figure 1.10).
Table 1.4| kinetic parameters for Thioredoxin from different organisms
Trx Disulfide substrate k
(M-1s-1)
Temp
(Cº)
pH Ref.
E.coli Trx1 Insulin 105 25 7 [113]
E.coli Trx1 (I27G32C-A75C)8 2.5x105 25 7.2 [98]
E.coli Trx2 (I27G32C-A75C)8 1.8x105 25 7.2 [98]
Human Trx2 (I27G32C-A75C)8 6.5x105 25 7.2 [98]
Human Trx1 (I27G32C-A75C)8 5.2x105 25 7.2 [98]
Human Trx1 hPrxII 2.1x105 25 7.4 [78]
Pea Trxm (I27G32C-A75C)8 2.8x105 25 7.2 [98]
P. falciparum Trx1 (I27G32C-A75C)8 4.3x105 25 7.2 [98]
Poplar Trx h3 (I27G32C-A75C)8 1.2x105 25 7.2 [98]
Poplar Trx h1 (I27G32C-A75C)8 2.2x105 25 7.2 [98]
(I27G32C-A75C)8 used as a substrate is a polyprotein composed of eight domains of the 27th module of human
cardiac titin in which each module contains an engineered disulfide bond between the 32nd and 75th positions
[114]. Table and legend adapted from ref. [98,114,115].
The Trx reducing mechanism thus seems to be the outcome of an evolutionary pressure to develop
an enzymatic process to reduce disulfide with rates constant that would have been not achievable
with simple chemical reagents.
Sulfiredoxins
Sulfiredoxin (Srx) is the enzyme responsible for the reduction of Prx-SO2-. It is conserved majorly amongst eukaryotes (Figure 1.11), in agreement with observations that prokaryotic Prx are less
sensible to hyperoxidation. Studies with the yeast Srx showed that the reduction requires also ATP
hydrolysis, Mg2+, and a thiol as an electron donor [44].
Figure 1.11| Relatedness tree for Sulfiredoxin sequences. An unrooted phylogenetic tree of 335 Srx
16
non-redundant protein database by BLAST on January 31, 2014, with an expect threshold of 100 using the human Srx1 sequence, and additional searches using distantly related Srx sequences did not identify further homologues. Sequences were aligned with MUSCLE, and evolutionary distances were calculated using PhyML. Figure and legend from ref [92]
The limiting step in this reaction is the formation of a thiosulfinate intermediate[45,89–91] (Srx-Prx) which existence has been confirmed for yeast [90] and human [116]. The resolution of this complex
may generate an intramolecular disulfide bond Srx-SS, that is then recovered by Trx or use
alternative pathways through GSH[117] (Figure 1.12).
Figure 1.12| Comparison of the proposed sulfinic acid reduction mechanism of Sulfiredoxin. Path 1
represents the mechanism originally proposed by Biteau et al. [44]. Path 2 incorporates modifications to the reaction pathway as suggested by Jeong et al. [117]. Step 1 involves the formation of the sulfinic acid phosphoryl ester intermediate. In Step 2 of the reaction, the addition of a thiol group leads to the formation of different thiosulfinate intermediates. This intermediate is subsequently resolved by GSH in Step 3. The resulting sulfenic acid form of Prx could then go on to react with Srx, GSH, and the resolving Cys of the adjacent Prx molecule to form Prx-SP–S-Srx, Prx-SP–S-G, Prx-SP–SR-Prx species. Figure and legend from ref
[118].
The reactivation is concentration dependent from Srx and limited by the Srx-Prx complex formation
[45,91,116]. The kcat values for the rat, human, and Arabidopsis thaliana Srx range from 0.1 to 1.8 min-1 [45,119–121].
The range of specific activities, 7–13 nmol min-1 mg-1 of protein (with hSrx ~10 nmol min-1mg-1), indicates that the Srx proteins are highly inefficient enzymes[121].
Alternative defense mechanism
Beyond Prx other enzymatic defenses have been developed by the organism to counterbalance
H2O2 increasing concentrations.
Glutathione peroxidase (GPx) catalyzes the reduction of H2O2 and organic peroxides by GSH. The reaction is coupled with NADPH oxidation, via the reduction of GSSG catalyzed by glutathione
reductase (GR). GPx has a homotetramer arrangement, in which every subunit contains a
selenocystein that gets oxidized to selenenic acid. This reacts consecutively with two GSH
molecules, the first forming a mixed selenenylsulfide, and the second reacting with this to produce
17
Catalase (Cat) is an enzyme typically located in peroxisomes that behaves as a dismutase at H2O2 concentrations above nanomolar and as a peroxidase at lower concentrations [124]. It is present in
almost all aerobic organisms and many anaerobic ones. The catalase cycle takes place in two
steps: the first H2O2 molecule oxidizes the heme to an oxyferryl species (compound I), the second H2O2 molecule is used as a reductant of compound I to regenerate catalase and release water and oxygen[125].
Hansen et al. [126] showed that the concentration of oxidizable protein thiols in human cell lines is in the order of 10 mM, which is comparable or higher than GSH concentrations. However, only a
small fraction of these thiols are very reactive[37], and none of these is sufficiently abundant to
contribute significantly for the H2O2 clearance capacity of the cells.
Hydrogen peroxide signal processing
The antioxidant enzymes (e.g. catalase, glutathione peroxidase, and the peroxiredoxins) maintain
endogenous cellular concentrations of H2O2 in the sub-micromolar range, and they show redox-sensitive transcription in order to increase their activities in response to oxidative insults [58,127].
Thiolate groups are expected to react with H2O2 at rate constants in a range of 18-26 M-1s-1 at 37ºC [128]. Other than those in the active centers of peroxidases and peroxiredoxins, few protein thiols
characterized to date have H2O2 reactivities above 100 M-1s-1 [129,130] (see Table 1.2), and none of these is sufficiently abundant to compete with the H2O2 clearance capacity of the cells.
Given the tight control of the H2O2 concentration and the low reactivities and expression of the possible signaling targets it is unlikely that a direct oxidation of these would be the main mechanism
of signal transmission. A possible solution to this conundrum, the “flood-gate hypothesis”, was proposed by Wood et al. [82]. In this hypothesis, Prxs act as a peroxide floodgate, controlling the oxidants concentration and protecting the most reactive thiols. The inactivation of Prx by
hyperoxidation caused by a local spike in the H2O2 concentration would allow H2O2 to temporarily accumulate and trigger the signaling cascade. This hypothesis relies on a high degree of
coordination between NOX and antioxidant defense for the propagation of the signal, and although
this may be possible in IOS conditions it seems unlikely under BOS/LOS and adds a potentially
slow tier in the signaling cascade, delaying the sensing process.
An alternative, but not exclusive, explanation of the peroxide signal propagation is the “redox-relay”. In this scenario highly reactive proteins as Prxs or GPxs mediate the transduction by sensing and
oxidizing specific protein thiols.
Here we focus on the capacity of the PTTRS to behave as a readout of the redox signals transferring
disulfide moiety to the target proteins avoiding direct interaction, and possible irreversible damage
The Peroxiredoxin/Thioredoxin/Thioredoxin Reductase System as hydrogen
peroxide sensor.
The conserved characteristics, even amongst kingdoms, and the high level of expression make of
the PTTRS a perfect candidate for mediating redox signals. In particular, it is becoming more
18
Recently Sobotta et al. ref. [131], reported that in HEK293T cells hPrxII directly oxidize the signaling protein STAT3 [131].
STAT3 is a member of the STAT protein family. Proteins in this family translocate to the cell nucleus
upon activation, and there they activate the STAT response (e.g. pro-oncogenic factor, interleukin-6 pathways and NF-kB)[132]. Specifically, STAT3 responds to ligands interferons, growth factors and Interleukin-6 and may be activated also via MAPK [132].
Another redox regulated protein, linked to inflammatory response and cancer is NF-kB. Reduced
Trx react with NF-kB allowing it to translocate in the nucleus and induce the response (i.e. IL-6 thus possibly activating STAT3)[133]. Important are also the evidences of redox relay that are activated
upon oxidative insults due to the oxidations through Trx[134,135].
The PTTR system components, in particular Prx and Trx, show also synergies in the regulation of
oxidative stress defense in a redox relay fashion by regulating the Pap1[95,136,137] and
Yap1[138,139] pathways respectively in S.pombe and S. cerevisiae.
García-Santamarina et al. ref. [135] showed that treatment of Schizosaccharomyces pombe (S. pombe) with 0.2 mM H2O2 (below the toxicity levels for this organism), induces a transient general oxidation of thiols and the consequent formation of disulfide in many proteins. These include
enzymes involved in antioxidant functions, Trx substrates, proteins related to proteasome,
ribosomal proteins and metabolic enzymes. The authors also found mixed disulfides of Trx1 with
target proteins in extracts of H2O2-treated cells. Such mixed disulfides are intermediates in the normal oxidation/reduction of protein thiols/disulfides by Trx. Subsequent studies from the same
author, ref. [134], showed that in S. pombeΔTrxR deletants the accumulated oxidized form of Trx
oxidizes a variety of thiol-containing proteins, and that these proteins are not oxidized in ΔTrxΔTrxR double mutant. Likewise, Baty et al. ref. [140] showed that treatment of Jurkat cells with H2O2 leads to a selective oxidation of the most reactive protein thiols. Altogether, these observations indicate
that oxidative pulses lead to a quick oxidation of the Trx pool and in turn trigger a substantial
oxidation of solvent-exposed protein thiols to disulfides.
PTTRS components also interact with different crucial proliferation/apoptosis regulatory factors.
Reduced Trx is able to form a complex with apoptosis signaling kinase-1 (Ask-1) inhibiting its
activation[141]. Upon oxidative stress Trx oxidation by hPrxI scavenging activity has been shown
to lead to the dissociation and activation of Ask-1[142,143] and eventually to cell death (Figure
1.13).
Figure 1.13| Trx-Ask1 interaction. The interaction between Trx and ASK1 is redox dependent and in turn