Polymorphism of locus DRB3.2 in populations
of Creole Cattle from Northern Mexico
Ilda G. Fernández
1,2, José Gonzalo Ríos Ramírez
2, Amanda Gayosso Vázquez
3,
Raúl Ulloa Arvizu
3and Rogelio A. Alonso Morales
31
Departamento de Ciencias Médico Veterinarias, Universidad Autónoma Agraria Antonio Narro,
Torreón, Coahuila, México.
2
Departamento de Reproducción, Facultad de Zootecnia, Universidad Autónoma de Chihuahua,
Chihuahua, Chihuahua, México.
3
Departamento de Genética y Bioestadística, Facultad de Medicina Veterinaria y Zootecnia,
Universidad Nacional Autónoma de México, México, D.F., México.
Abstract
The polymorphism of locus BoLA-DRB3.2 of the Major Histocompatibility Complex was evaluated in two northern Mexican Creole cattle populations, Chihuahua (n = 47) and Tamaulipas (n = 51). The BoLA-DRB3.2 locus was typed by amplification and digestion with restriction endonuclease enzymes (PCR-RFLP). Fifty-two alleles were detected (28 previously reported and 24 new ones). In the Chihuahua population, 18 alleles and 5.5 effective alleles were found, while in the Tamaulipas population there were 34 and 10.8, respectively. The allele frequencies ranged from 0.011 to 0.383 in Chihuahua and from 0.010 to 0.206 in Tamaulipas. The frequencies of the new alleles in both cattle populations were low (0.010 to 0.053). The expected heterozygosity was 0.827 and 0.916, respectively, for the Chi-huahua and Tamaulipas populations. Both populations presented a heterozygote deficit: [ChiChi-huahua FIS= 0.1 (p = 0.019) and Tamaulipas FIS= 0.317 (p < 0.001)]. In conclusion, this study showed that the Mexican Creole cattle have many low-frequency alleles, several of which are exclusive to these populations. Genetic distances obtained show that the Mexican Creole cattle population is composed of independent populations, far apart from other South Ameri-can Creole populations.
Key words:BoLA-DRB3.2, polymorphism, Creole cattle, PCR-RFLP.
Received: November 30, 2007; Accepted: June 30, 2008.
Introduction
The Mexican Creole cattle originated from Bos tauruspopulations brought to Mexico by the Spaniards in the XVI century, mainly from the Iberian peninsula. Cur-rently, the Mexican Creole cattle population consists of un-selected populations with low productivity features, such as low body weight and low milk production. On the other hand, they seem to have the ability of surviving in extreme climates and of resisting to diseases. These cattle are seri-ously endangered by the rapid displacement due to im-proved modern breeds, but some populations still remain in remote areas in South, Central and North Mexico. In north-ern Mexico, Creole cattle are raised by small communities under precarious extensive conditions (input and low-management) in a semi-arid climate. In the state of
Chihua-hua, they are reared by peasants and by indigenous breeders of the Sierra Tarahumara, ever since their original introduc-tion by Jesuit priests in 1667 (Ríos, 2004). In the state of Tamaulipas, similarly, the Creole cattle are raised by small farmers, although the breed was introduced from different regions of the country, mainly by a genetic rescue program that the Mexican government started around 1980.
Certain populations of Mexican Creole cattle have become a biotype of commercial interest for rodeo sporting events in the USA and Canada, because they possess fea-tures required for this activity. These feafea-tures are hardiness, docility, speed, as well as tolerance to physical punishment and orientation, shape, length, thickness, and strength of the horns (Félix Portilloet al., 2006).
One of the characteristics that have probably evolved due to natural selection is the resistance to infectious dis-eases, because the Creole cattle are usually not immunized. The Major Histocompatibility Complex (MHC) is one of the main components of the immune system. Also called bovine lymphocyte antigen (BoLA) complex in cattle, it is www.sbg.org.br
Send correspondence to Ilda G. Fernández. Departamento de Ciencias Médico Veterinarias, Universidad Autónoma Agraria An-tonio Narro, Periférico Raúl López Sánchez y Carretera a Santa Fe, 27054 Torreón, Coahuila, México. E-mail: ilda_fernandez_ [email protected].
located on chromosome 23 and contains some genes in-volved in antigen presentation necessary to trigger the im-mune response (Hesset al., 1999). The class II BoLA-DR region is composed of one DRA locus and at least three DRB loci (DP, DQ, and DR) (Muggli-Cockett and Stone, 1989). The MHC allele diversity is associated with the abil-ity to recognize a large number of antigens, resulting in a more efficient immune response (Behlet al., 2007). So it may be interesting to assess the level of allele diversity in the BoLA-DRB3.2 gene in a population that is under great pressure for survival under tough conditions, such as the Mexican Creole cattle populations. In addition, this poly-morphism can be used to study the genetic relationships be-tween populations and to assess their levels of genetic differentiation. Presently, extensive information is avail-able on the levels of genetic diversity of exon 2 of the DRB3 gene in different populations of cattle, obtained by amplification of this segment by PCR, and subsequent di-gestion with endonucleases (Gilliespieet al., 1999; Kellyet al., 2003; Martinezet al., 2005; Behlet al., 2007). The ob-jective of this study was to investigate the level of genetic diversity present in the MHC BoLA-DRB3.2 locus in two Creole cattle populations from northern Mexico.
Materials and Methods
Samples were obtained from 98 Creole Bos taurus
cattle from two northern states of Mexico: Chihuahua State samples were obtained from three localities (Morelos: n = 26, Chinipas: n = 7, and Guachochi: n = 14), all located in Sierra Tarahumara. All of the Tamaulipas samples came from a particular region (Soto la Marina; n = 51).
Amplification of BoLA-DRB3
DNA was purified from blood samples following standard procedures (Sambrook et al., 1989). A 284-bp segment of DNA containing the BoLA-DRB3.2 gene was amplified by PCR. For this purpose, we used a pair of oligonucleotides described previously by van Eijk et al.
(1992) (HL030: 5’-ATCCTCTCTCTGCAGCACATTTC C-3’ and HL032: 5’-TCGCCGCTGCACAGTGAAACTC TC-3’). The amplification of the BoLA-DRB3.2 gene was performed in a single PCR reaction rather than in two sub-sequent hemi-nested reactions as originally proposed by van Eijket al.(1992). PCR reactions were carried out in a total volume of 50µL, containing 150 ng of genomic DNA,
1.5 mM MgCl2, 10 mM Tris – HCl pH 8.4, 50 mM KCl,
1 mg/mL gelatin, 0.15 mg/mL bovine serum albumin, 0.1% Triton-X100, 0.2 mM deoxynucleotide triphosphates (dNTPs; Biogénica S.A. de C.V. Mexico), 1µM of each
oligonucleotide HL030 and HL032, and 2.5 U Taq DNA polymerase (Biogénica, S. A. de C. V. Mexico). Amplifica-tion was carried out in a Thermal Cycler (Omn-E HYBAID, UK), with an initial denaturation step of 3 min at 94 °C, followed by 30 cycles of 30 s at 94 °C, 30 s at 68 °C
and 30 s at 72 °C, and a final extension step of 5 min at 72 °C.
PCR-RFLP analysis
Three aliquots of 60µL were digested independently,
each containing 10µL of the PCR-amplified products, 1X
of the recommended digestion buffer, and 5 U of each of the restriction enzymes RsaI, BstYI, and HaeIII (New Eng-land Biolabs Inc., Mass., USA), and incubated at the rec-ommended temperature for 14 h. As a control of BstYI complete digestion, 10µL of the digestion mix were added
to 160 ng of pBR322 DNA and incubated in parallel.
Polyacrylamide gel electrophoresis
The digested products were resolved by vertical gel electrophoresis (8 x 10 cm) on an 8% polyacrylamide gel (Sambrooket al., 1989), and stained with silver nitrate us-ing a commercial kit (BIO-RAD Silver Stain Plus 161-0448). As a molecular weight marker, pBR322 DNA digested with MspI was used. The size of the digestion products was estimated with the RFLP Molecular Weight Program, version 2.17 (UVP, Inc., Upland, CA, USA). Al-lele identification from the restriction patterns and its no-menclature have been previously reported by van Eijket al.
(1992), Gelhauset al. (1995), Maillardet al.(1999), and Gilliespieet al.(1999).
Definition of alleles in heterozygote individuals
The PCR products were cloned into a T vector (pUC19 DNA was digested with EcoRV and T tails added at the 3’ end, as described by Sambrooket al., 1989). The cloned products were subjected to PCR and the amplified products digested with restriction enzymes.
Statistical analysis
Statistical analysis was carried out using the GENEPOP v.3.4 program (Raymond and Rousset, 1995a) to calculate allele frequencies, the value FIS (Weir and
gene diversity was calculated from the previously reported frequencies (Nei and Chesser, 1983), and the effective al-lele number (ne) was obtained as the inverse of the
homo-zygosity (Nei, 1987).
Results
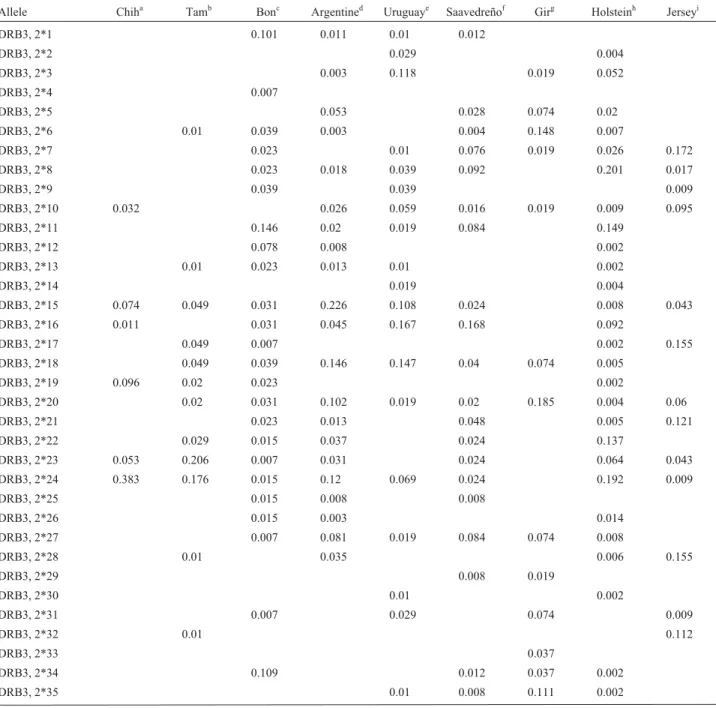
In the 98 animals tested, we identified 52 BoLA-DRB3.2 alleles; 28 had already been reported (van Eijket al.,1992; Gelhaus et al.,1995; Maillard et al.,1999; Gilliespieet al.,1999; Behlet al., 2007) and 24 were new patterns. The identified alleles and their frequencies are shown in Table 1. In the Chihuahua Creole cattle, we found
18 alleles of which seven were new, and in the Tamaulipas Creole cattle we found 34 alleles, of which 17 were new; these two populations shared seven alleles, of which two were new ones (nbd and sba).
Genetic diversity was 0.83 and 0.92 for Chihuahua and Tamaulipas Creole cattle, respectively. The effective allele number was 5.5 for Chihuahua and 10.8 for Tamaulipas. As shown in Table 1, in the Chihuahua popu-lation, the cumulative frequency of six alleles out of 18 (*24, *19, ibb, *15, xbb, and *23) accounted for 75% of the allele frequency. For the population of Tamaulipas, eight alleles out of 34 (*23, *24, *39, *17, *18, *15, tbb, and *22) accounted for 66.6% of the whole allele frequency.
Table 1- Allele distribution and gene frequency of locus DRB3.2 in Creole cattle and in improved breeds.
Allele Chiha Tamb Bonc Argentined Uruguaye Saavedreñof Girg Holsteinh Jerseyi
DRB3, 2*1 0.101 0.011 0.01 0.012
DRB3, 2*2 0.029 0.004
DRB3, 2*3 0.003 0.118 0.019 0.052
DRB3, 2*4 0.007
DRB3, 2*5 0.053 0.028 0.074 0.02
DRB3, 2*6 0.01 0.039 0.003 0.004 0.148 0.007
DRB3, 2*7 0.023 0.01 0.076 0.019 0.026 0.172
DRB3, 2*8 0.023 0.018 0.039 0.092 0.201 0.017
DRB3, 2*9 0.039 0.039 0.009
DRB3, 2*10 0.032 0.026 0.059 0.016 0.019 0.009 0.095
DRB3, 2*11 0.146 0.02 0.019 0.084 0.149
DRB3, 2*12 0.078 0.008 0.002
DRB3, 2*13 0.01 0.023 0.013 0.01 0.002
DRB3, 2*14 0.019 0.004
DRB3, 2*15 0.074 0.049 0.031 0.226 0.108 0.024 0.008 0.043
DRB3, 2*16 0.011 0.031 0.045 0.167 0.168 0.092
DRB3, 2*17 0.049 0.007 0.002 0.155
DRB3, 2*18 0.049 0.039 0.146 0.147 0.04 0.074 0.005
DRB3, 2*19 0.096 0.02 0.023 0.002
DRB3, 2*20 0.02 0.031 0.102 0.019 0.02 0.185 0.004 0.06
DRB3, 2*21 0.023 0.013 0.048 0.005 0.121
DRB3, 2*22 0.029 0.015 0.037 0.024 0.137
DRB3, 2*23 0.053 0.206 0.007 0.031 0.024 0.064 0.043
DRB3, 2*24 0.383 0.176 0.015 0.12 0.069 0.024 0.192 0.009
DRB3, 2*25 0.015 0.008 0.008
DRB3, 2*26 0.015 0.003 0.014
DRB3, 2*27 0.007 0.081 0.019 0.084 0.074 0.008
DRB3, 2*28 0.01 0.035 0.006 0.155
DRB3, 2*29 0.008 0.019
DRB3, 2*30 0.01 0.002
DRB3, 2*31 0.007 0.029 0.074 0.009
DRB3, 2*32 0.01 0.112
DRB3, 2*33 0.037
DRB3, 2*34 0.109 0.012 0.037 0.002
Table 1 (cont.)
Allele Chiha Tamb Bonc Argentined Uruguaye Saavedreñof Girg Holsteinh Jerseyi
DRB3, 2*36 0.049 0.116
DRB3, 2*37 0.007 0.08
DRB3, 2*38
DRB3, 2*39 0.069
DRB3, 2*40 0.02 0.007
DRB3, 2*41 0.031
DRB3, 2*42 0.056
DRB3, 2*43
DRB3, 2*44 0.023 0.010 0.019
DRB3, 2*45 0.021 0.010
DRB3, 2*46 0.023
DRB3, 2*47 0.031 0.037
DRB3, 2*48 0.015
DRB3, 2*49
DRB3, 2*50 0.011 0.010
DRB3, 2*51
DRB3, 2*53 0.010
ibd 0.032
jbb 0.021
nbd 0.021 0.010
seb 0.011
taa 0.010
tbb 0.039
tbd 0.010
ubb 0.010
ubd 0.010
vbd 0.020
xbb** 0.053
dba 0.020
dbb 0.010
fbb* 0.032
gbb 0.021
ibb* 0.096 0.020
ibe 0.011
kbd 0.010
mab 0.010
mbb 0.010
naa 0.020
gba 0.010
nbe 0.020
obd 0.010
sba 0.021 0.010
n 47 51 162 194 51 125 28 835 66
Genetic diversity 0.827 0.916 0.939 0.920 0.916 0.915 0.916 0.865 0.885
ne 5.5 10.8 15.6 8.8 10.8 11.8 10.0 7.4 8.2
Alleles total 18 34 31 11 15 22 22 27 13
ne= Effective number of alleles. Alleles 1-53 ISAG (2002). n = Sample size.aChihuahua,bTamaulipas,cBlancorejinegro; Martínezet al.(2005), d
The FISparameter assesses the deviation between
ex-pected and observed heterozygosity (Weir and Cockerham, 1984). In our study, the FISvalues obtained for the
Chihua-hua Creole and Tamaulipas populations were 0.1 (p = 0.0196) and 0.317 (p < 0.001), respectively. Positive values mean that both groups present a heterozygote defi-cit. The FISvalues for the three subpopulations of the
Chi-huahua population (Chinipas, Guachochi and Morelos) were also estimated (-0.063, 0.233 and 0.075, respec-tively), and only the Morelos population presented hetero-zygote deficiency at a significant level (p = 0.034).
The FSTvalue measures the degree of genetic
differ-entiation between populations. This value was FST= 0.043
between the Chihuahua and Tamaulipas groups, which means that 4% of the genetic variation corresponded to dif-ferences between populations, whereas the remaining 96% were due to differences among individuals within each population. These results indicate little genetic differentia-tion between these two populadifferentia-tions. We also performed a genetic differentiation test among the subpopulations of Chihuahua (Morelos, Chinipas and Guachochi) and Ta-maulipas (FST= 0.033). The allele distribution was similar
among the three Chihuahua subpopulations (p > 0.05) and between Chinipas and Tamaulipas (p > 0.05); however, it was different between Guachochi and Morelosvs. Tamau-lipas (p < 0.001). In order to analyze the genetic relation-ships of the Mexican Creole populations with other American Creole cattle and with other commercial breed cattle, we evaluated their genetic distances by the Reynolds method, using the allele frequencies reported for Argentina (Giovambattistaet al., 1996), Uruguay (Kellyet al., 2003), Colombia (Martinez et al., 2005), Bolivia (Ripoliet al., 2004), Holstein and Jersey breeds (Sharifet al., 1998), and for the non-taurine population Zebu Gir (da Motaet al., 2002). The genetic relationships among the different popu-lations are shown in Figure 1.
Discussion
The results of this study show that Chihuahua and Tamaulipas Creole cattle have a high degree of BoLA-DRB3.2 polymorphism, in concordance with findings in other populations, such as the American Creole cattle (Gio-vambattistaet al., 1996; Kellyet al., 2003; Ripoliet al., 2004; Martinezet al., 2005), European taurine breeds (Sha-rifet al., 1998), Asian taurine breeds (Takeshima et al., 2003), and the Zebu Gir breed (da Motaet al., 2002; Behlet al., 2007). In this study, allele DRB3.2*24 showed the highest frequency (0.38) in Chihuahua Creole cattle, and it has also been reported to be present at frequencies higher than 0.1 in all taurine populations (Giovambattistaet al., 1996; Sharifet al., 1998; Gilliespieet al., 1999; Kellyet al., 2003). This allele was further reported to be associated with a higher antibody-mediated immune response and with a lower cell-mediated immune response (Ruppet al., 2007). In the Tamaulipas population, the most frequently found al-lele was DRB3.2*23 (0.206). This alal-lele has been detected in almost all populations of taurine cattle (Giovambattista
et al., 1996; Sharifet al., 1998; Gilliespieet al., 1999), but not in Uruguayan Creole cattle (Kelly et al., 2003). The high frequency of alleles DRB3.2*23 and DRB3.2*24 found in Tamaulipas and Chihuahua Creole cattle may be the result of positive natural selection, but this remains to be demonstrated.
Allele DRB3.2*10 was observed at low frequencies (< 0.1) in most of the groups except in Tamaulipas Creole and Blancorejinegro Creole (Martinezet al., 2005). Allele DRB3.2*16 was more frequent in Uruguayan Creole (Kellyet al., 2003) and Saavedreño (Ripoliet al., 2004), but in our study it was not detected in Tamaulipas Creole and was present in Chihuahua Creole at a low frequency (1.1%). Gelhauset al.(1995) demonstrated the presence of eight alleles in African breeds (N’Dama and Boran); we found some of these alleles in the Tamaulipas population (DRB3.2*16 and DRB3.2*32) and in the Chihuahua Creole cattle (DRB3.2*6 and DRB3.2*10).
It is known that the MHC genes are highly polymor-phic in the populations studied so far (Dietzet al., 1997). Although we found a high number of alleles in Mexican populations, the Chihuahua Creole cattle had the lowest ef-fective allele number (5.5), even lower than that of Holstein (7.4) and Jersey (8.2) populations, which are known to be undergoing intense selection and inbreeding. Actually, in the Mexican Creole cattle many alleles at low frequencies were found, but presenting some exclusiveness. In fact, considering the ratio between the effective allele number and the total number of alleles, the Argentine and Uru-guayan Creole showed a high relationship (0.8 and 0.72, respectively), whereas in the Chihuahua and Tamaulipas Creole cattle this relationship was very low (0.3 and 0.317, respectively). This may be the result of natural selection on a particular BoLA allele pool, leading to the development of resistance to local infectious diseases.
Of the novel alleles reported in this study, sequence analysis was made in 10 of them by Félix Portilloet al.
(2006), who found that two of these corresponded to the previously reported alleles DRB3*1602 and DRB3*1501. The remaining eight alleles rendered unique nucleic acid sequences after a search in the official BoLA database. Even though the nucleotide sequencing of BoLA genes is a more precise way of allele identification and comparison, for laboratories with limited resources the PCR-RFLP tech-nique is a more useful and effective tool for BoLA-DRB3.2 genotyping, because it is easier to perform, cheaper and faster.
In the Chihuahua and Tamaulipas Creole cattle, we observed an excess of homozygosity that may be due to the combined effect of inbreeding and the Wahlund effect. The Chihuahua Creole cattle animals belonged to three sub-populations from the Sierra Tarahumara, which are isolated and have only few sires available. In turn, the Tamaulipas Creole cattle, although the population descended from ani-mals from different regions of Mexico, are currently con-fined and consist of small, closed herds.
Finally, Figure 1 presents a dendrogram that summa-rizes the genetic distances and shows that the Mexican Creole cattle is an independent branch, far apart from the South American Creole populations. This may indicate that this population has a different origin from those popula-tions, or that it has experienced extreme genetic differentia-tion, probably due to a founder effect. It is also evident that there is extensive differentiation among the different popu-lations of Creole cattle in America, although the Argentin-ean and Uruguayan populations are closer related, being placed in a common node. Furthermore, although the Chi-huahua and Tamaulipas Creole populations are in the same branch, there is a great genetic distance between them. This may be the result of the ancient isolation and independent origin of the Chihuahua group. This information is impor-tant for the knowledge of this endangered genetic resource and for the establishment of strategic conservation pro-grams. Furthermore, our findings suggest that these local cattle populations may be a valuable reservoir of genetic di-versity for the development of defined local breeds or in the improvement of commercial cattle lines.
Acknowledgments
The authors wish to express their gratitude to all members of the Laboratorio de Genética Molecular, Facul-tad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México, for their help, and particu-larly to MC María del Carmen Castro Méndez and to Dr. Refugio Cortés Fernández.
References
Behl JD, Verma NK, Behl R, Mukesh M and Ahlawat SPS (2007) Characterization of genetic polymorphism of the bovine
lymphocyte antigen DRB3.2 locus in Kankrej cattle (Bos indicus). J Dairy Sci 90:2997-3001.
da Mota AF, Gabriel JE, Martinez ML and Coutinho LL (2002) Distribution of bovine lymphocyte antigen (BoLA-DRB3) alleles in Brazilian dairy Gir cattle (Bos indicus). Eur J Immunogenet 29:223-227.
Dietz AB, Cohen ND, Timms L and Kehrli Jr ME (1997) Bovine lymphocyte antigen class II alleles as risk factors for high somatic cell counts in milk of lactating dairy cows. J Dairy Sci 80:406-412.
Félix Portillo M, Ríos Ramírez JG, Erosa de la Vega GE and Rodríguez Almeida F (2006) Sequencing of new BoLA-DRB3.2 alleles detected in Mexican Creole cattle. Téc Pecu Méx 44:15-25.
Felsenstein J (1993) PHYLIP: Phylogeny Inference Package, v. 3.55. University of Washington, Seattle.
Gelhaus A, Schnittger L, Mehlitz D, Horstmann RD and Meyer CG (1995) Sequence and PCR-RFLP analysis of 14 novel BoLA-DRB3 alleles. Anim Genet 26:147-153.
Gilliespie BE, Jayarao BM, Dowlen HH and Oliver SP (1999) Analysis and frequency of bovine lymphocyte antigen DRB3.2 alleles in Jersey cows. J Dairy Sci 82:2049-2053. Giovambattista G, Golijow CD, Dulout FN and Lojo MM (1996)
Gene frequencies of DRB3.2 locus of Argentine Creole cat-tle. Anim Genet 27:55.
Guo SW and Thompson EA (1992) Performing the exact test of Hardy-Weinberg proportions for multiple alleles. Biome-trics 48:361-372.
Hess M, Goldammer T, Gelhaus A, Ried K, Rappold G, Eggen A, Bishop MD, Schwerin M and Horstmann RD (1999) Physi-cal assignment of the bovine MHC class IIa and IIb genes. Cytogenet Cell Genet 85:244-247.
Huson D and Bryant D (2006) Application of phylogenetic net-works in evolutionary studies. Mol Biol Evol 23:254-267. Kelly L, Nicolini P, D’Angelo M, Nimo A, Rincón G, Piagio J and
Postiglioni A (2003) Polimorfismo de gen DRB3.2 en bovi-nos criollos del Uruguay. Arch Zootec 52:77-80.
Maillard JC, Renard C, Chardon P, Chantal I and Bensaid A (1999) Characterization of 18 new BoLA-DRB3 alleles. Anim Genet 30:200-203.
Martínez R, Toro R, Montoya F, Burbano M, Tobón J, Gallego J and Ariza F (2005) Caracterización del locus BoLa-DRB3 en ganado criollo colombiano y asociación con resistencia a enfermedades. Arch Zootec 54:349-356.
Muggli-Cockett NE and Stone RT (1989) Partial nucleotide se-quence of a bovine major histocompatibility class II DR beta-like gene. Anim Genet 20:361-367.
Nei M (1987) Molecular Evolutionary Genetics. Columbia Uni-versity Press, New York, 512 pp.
Nei M and Chesser K (1983) Estimation of fixation indices and gene diversities. Ann Hum Genet 47:253.
Raymond M and Rousset F (1995a) GENEPOP, v. 1.2: Popula-tion genetics software for exact tests and ecumenicism. J Hered 86:248-249.
Raymond M and Rousset F (1995b) An exact test for population differentiation. Evolution 49:1280-1283.
Reynolds J, Weir BS and Cockerham CC (1983) Estimation of the coancestry coefficient: Basis for a short-term genetic dis-tance. Genetics 105:767-779.
Ripoli MV, Lirón VJP, De Luca JC, Rojas F and Giovambattista G (2004) Gene frequency distribution of the BoLA-DRB3 locus in Saavedreño Creole dairy cattle. Biochem Genet 42:231-240.
Rousset F and Raymond M (1995) Testing heterozygote excess and deficiency. Genetics 140:1413-1419.
Rupp R, Hernandez A and Mallard BA (2007) Association of bo-vine leukocyte antigen (BoLA) DRB3.2 with immune re-sponse mastitis, and production and type traits in Canadian Holstein. J Dairy Sci 90:1029-1038.
Sambrook J, Fritsch EF and Maniatis T (1989) Molecular Clon-ing: A Laboratory Manual. 2nd edition. Cold Spring Harbor Press, New York.
Sharif S, Mallard BA, Wilkie BN, Sargeant JM, Scott HM, Dek-kers JCM and Leslie KE (1998) Associations of the bovine major histocompatibility complex DRB3 (BoLA-DRB3) al-leles with occurrence of diseases and milk somatic cell score in Canadian dairy cattle. Anim Genet 29:185-193.
Takeshima S, Saitou N, Morita M, Inoko H and Aida Y (2003) The diversity of bovine MHC class II DRB3 genes in Japa-nese Black, JapaJapa-nese Shorthorn, Jersey and Holstein cattle in Japan. Gene 316:111-118.
van Eijk MJT, Stewart-Haynes JA and Lewin HA (1992) Exten-sive polymorphisms of the BoLA-DRB3 gene distinguished by PCR-RFLP. Anim Genet 23:483-496.
Weir B and Cockerham C (1984) Estimating F-statistics for the analysis of population structure. Evolution 38:1358-1370.
Internet Resources
Report of the 8th Workshop of the ISAG BoLA Nomenclature Committee (2002). http://www.projects.roslin.ac.uk/bola/ wk98.html (June 2, 2007).
Associate Editor: Luiz Lehmann Coutinho