Genetic diversity of cultivated accessions and wild species of rubber tree using EST‑SSR markers
Texto
Imagem

Documentos relacionados
Thermal insulation helps in preserving energy efficiency, which depends on the air temperature gradient (the difference between inside and outside of building), geographical
(2012) a inclusão crescente do calcário pedrisco na ração de poedeiras em final de produção influenciaram negativamente as características de desempenho das aves
In the research described in this paper we used SSR markers covering the seven barley SSR linkage groups (Liu et al. , 1996) to investigate the genetic diversity and geo-
Através do planeamento deste Projecto de Parceria entre a ONGD Estímulo e a Rede de Farmácias Portuguesas pretende demonstrar-se que existem inúmeros objectivos que não
The results, that are based on the application of a questionnaire, show that teachers, despite their different academic backgrounds (all of them with a background
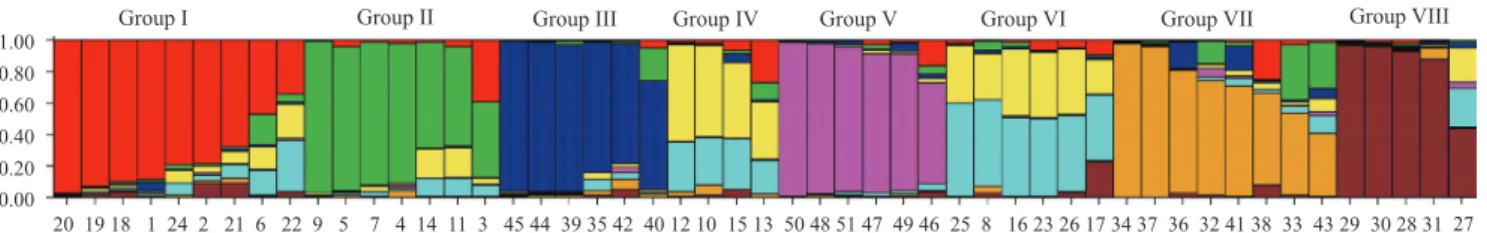
index ( θ ˆ) evaluated in 22 rubber tree genotypes (clones) and a control selected in the Brazilian Hevea breeding program.. (1) RYT, mean dry rubber yield in the third year
Our data showed that EST-SSR markers may be useful in coffee breeding programs, particularly for crossings orientation, diversity studies, genetic introgression and genetic
psicológica e social, a instituição de ensino deixa a desejar, pois não dispõe desses profissionais na área para dar um apoio aos alunos quando se fizer
