REVISTA
BRASILEIRA
DE
ANESTESIOLOGIA
OfficialPublicationoftheBrazilianSocietyofAnesthesiologywww.sba.com.br
REVIEW
ARTICLE
Satellite
glial
cells
in
sensory
ganglia:
its
role
in
pain
夽
Filipa
Alexandra
Leite
Costa
a,
Fani
Lourenc
¸a
Moreira
Neto
b,c,∗aFaculdadedeMedicina,UniversidadedoPorto,Porto,Portugal
bDepartmentofExperimentalBiology,CentrodeInvestigac¸ãoMédicadaFaculdadedeMedicinadoPorto(CIM-FMUP),
UniversidadedoPorto,Porto,Portugal
cNervousSystemMorphophysiologyGroup,InstitutodeBiologiaMoleculareCelular(IBMC),UniversidadedoPorto,
Porto,Portugal
Received30October2012;accepted15July2013 Availableonline6November2014
KEYWORDS
Satelliteglialcells; Sensoryganglia; Pain;
Intraganglionar communication; Purinergicreceptors
Abstract
Backgroundandobjectives: Satellite glial cells in sensory ganglia are a recent subject of researchinthefieldofpain andapossibletherapeutic targetinthefuture.Therefore,the aim ofthisstudy was tosummarize someoftheimportantphysiological andmorphological characteristicsofthesecellsandgatherthemostrelevantscientificevidenceaboutitspossible roleinthedevelopmentofchronicpain.
Content: Inthesensoryganglia,eachneuronalbodyissurroundedbysatelliteglialcells form-ingdistinctfunctionalunits.Thiscloserelationshipenables bidirectionalcommunicationvia aparacrinesignalingbetweenthosetwocelltypes.Thereisagrowingbodyofevidencethat glialsatellitecellsundergostructuralandbiochemicalchangesafternerveinjury,which influ-enceneuronalexcitabilityandconsequentlythedevelopmentand/ormaintenanceofpainin differentanimalmodelsofchronicpain.
Conclusions: Satelliteglialcellsareimportantintheestablishmentofphysiologicalpain,in additiontobeingapotentialtargetforthedevelopmentofnewpaintreatments.
© 2014SociedadeBrasileirade Anestesiologia.Publishedby ElsevierEditoraLtda.Allrights reserved.
PALAVRAS-CHAVE
Célulasgliaissatélite; Gângliosensitivo; Dor;
Célulasgliaissatélitedegângliossensitivos:oseupapelnador
Resumo
Justificativaeobjetivos: Ascélulasgliaissatélitedegângliossensitivossãoumobjetorecente depesquisanaáreadadoreumpossívelalvoterapêuticonofuturo.Assim,estetrabalhotem
夽 DepartmentofExperimentalBiology,CentrodeInvestigac¸ãoMédicadaFaculdadedeMedicinadoPorto(CIM-FMUP),Universidadedo
Porto,Portugal.
∗Correspondingauthor.
E-mail:[email protected](F.L.MoreiraNeto). http://dx.doi.org/10.1016/j.bjane.2013.07.013
Comunicac¸ão intraganglionar; Receptores purinérgicos
comoobjetivoresumiralgumasdascaracterísticasmorfológicasefisiológicasmaisimportantes destascélulasereunirasevidênciascientíficasmaisrelevantesacercadoseupossívelpapelno desenvolvimentodadorcrônica.
Conteúdo: Nosgângliossensitivoscadacorponeuronialéenvolvidoporcélulasgliaissatélite, formandounidadesfuncionaisdistintas.Estaíntimarelac¸ãopossibilitaacomunicac¸ão bidire-cional,atravésdeumasinalizac¸ãoparácrina,entreestesdoistiposdecélulas.Existeumnúmero crescentedeevidênciasdequeascélulasgliaissatélitesofremalterac¸õesestruturaise bio-químicas,apóslesãonervosa,queinfluenciamaexcitabilidadeneuronialeconsequentemente odesenvolvimentoe/oumanutenc¸ãodador,emdiferentesmodelosanimaisdedorcrônica. Conclusões:Ascélulasgliaissatélitesãoimportantesnoestabelecimentodadornãofisiológica econstituemumalvopotencialparaodesenvolvimentodenovostratamentosdador. ©2014SociedadeBrasileiradeAnestesiologia.PublicadoporElsevierEditoraLtda.Todosos direitosreservados.
Introduction
Pain has a physiological protective role acting as a war-ning signal to any threat to the physical integrity of the body,butitcanbecomeadiseaseinitselfwhenitpersists andrecursformorethan threemonths,becomingchronic anddevoid ofany biological function.1 Itis a perceptive,
complex, subjective, and multidimensional phenomenon. Differentformsof painareproducedby multiple molecu-larandcellularmechanismsactingaloneorincombination withthecentralandperipheralnervoussystems.2Insearch
ofnewtherapeutictargetsitisessentialtounderstandthe mechanismsresponsibleforthegenerationandmaintenance ofpainandidentifythecellsand/ormoleculesinvolved.3In
thiscontext, theglialcells of thecentral nervoussystem (CNS),andmorerecentlythoseofthesensoryganglia,have showntobeimportantinthesemechanisms,astheyhave theabilitytocommunicatewithneuronsandmodulateits activity.4,5 In sensory ganglia,particularly in dorsal spinal
ganglia(DSG) and trigeminalganglion (TG),satellite glial cells(SGC)establishaprivilegedrelationshipwiththe sur-roundingneuronal bodies.6 Interactions between SGCand
neuronsanditsconsequencesonneuronalexcitability are one of the most recent research focuses on the field of pain,asinthelasttenyearsthenumberofarticlesonthe rolethesecellsplayinneuronalactivityhasincreased sig-nificantly. Therefore,the aim of this study wastogather knowledgeaboutthemorphologicalandfunctional charac-teristicsofSGCanditsinteractionwiththeprimaryafferent neurons.Wealsoreviewedtheavailableinformationonthe changesseeninthesecellsindifferentmodelsforthestudy ofpaininanimalsanditsimpactonneuronalactivityand consequentlyinchronicpain.
The
SGC
The SGC,as well as theSchwann cells, arederived from thepluripotent neural crest cells.7 Morphologically, these
cellsarecharacterizedbyalaminar,irregularshape,usually
mononuclear, withlamellarexpansions andmicrovilli that increaseitssurfacearea.8---10Thesecellssurroundeach
neu-ronandtheproximalportionofitsaxon,formingasheath aroundeachcellbody.Eachcellbodysurroundedbyasheath of SGC forms a morphologically and functionally distinct unit.6,11 TheSGCfromasheathareconnectedtogetherby
adhesive and gap junctions, and are separated from the neighboring perineuronalsheath byconnectivetissue.12---14
Physiologically, SGC are regarded as the equivalent cells in the peripheral nervous system to the CNS astrocytes, withtheinvestigationof itsfeaturesmarkedbythis anal-ogy. Properties suchas regulation of ion concentration of extracellularspaceandthe recyclingofneurotransmitters aresharedwiththem.Theyaremolecularmarkersof glu-taminesynthase(GS),proteinsoftheS100familyinvolved in the regulation of intracellular calcium and expression of glial fibrillary acidic protein (GFAP).6
Electrophysio-logically, glial cells exhibit a highly negative membrane potentialatrestandexpressvoltage-dependentcalciumand potassium channels Kir4.1.15---17 They also express
numer-ous receptorsof bioactive molecules, potentiallyinvolved ininteractionswithothercells,withmanyofthemrecently implicated in the genesis and maintenance of chronic pain,namely,theP2Y18,19 andP2X
720 purinergic,calcitonin
gene-relatedpeptide(CGRP),21substanceP,22 andcytokine
and chemokine receptors----examples of which are tumor necrosis factor alpha (TNF␣)23 and interleukin-1beta
(IL-1)24andendothelin-B25and
N-methyl-d-aspartate(NMDAr)
receptors.26
Intraganglionar
communication
peculiarities encountered is the presence of several neu-rotransmitterreceptors inthecellbody,although thereis virtually no contactin synaptic ganglion.27
Electrophysio-logicalstudiesinvivo havefoundotherindicators showing that the excitation of DSG neurons led to the develop-mentofactionpotentialinneighboringneurons,aproperty called cross-excitation. This potential was confirmed in studies in vitro in which repeated stimulation of these neurons induced atransient depolarizationof neighboring neurons in this ganglion, probably mediated by chemical messengers.28,29Accordingtothisassumption,itwasfound
that in response to an electrical or chemical stimulation there is a calcium-dependent somatic release30 of
dif-fusiblechemicalmediatorscapableofalteringthesomatic excitabilityinsensoryganglia.Examplesofsuchmediators are substance P, adenosine triphosphate (ATP), gamma
-amino-butyricacid(GABA),CGRP,andglutamate.20,21,30---34
On the other hand, the cellular body is completely envelopedbytheGSCsheath,suggestingthattheeffectof thesemediatorsonsurroundingneuronsisindirect, involv-ing the SGC.34 The SGC peculiar arrangement in sensory
gangliaguaranteesanintimateassociationoftheneuronal bodywiththeSGC,allowingtheseglialcellstocontrolthe perineuralenvironmentandfacilitatenon-synaptic commu-nicationbetweenthesetwocelltypes.21,22,34,35 Indeed,the
existence of two-way interactions between sensory neu-rons and SGC was recently demonstrated.34,35 The way
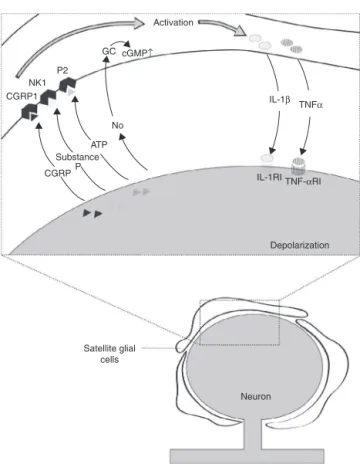
theCGS-neuron communication takesplace, the actorsin theprocess,anditsimpactonthe modulationofafferent informationarefarfrombeingunderstood.However,some potentialcandidatetomediatethisparacrinesignalingare substanceP,CGRP,cytokines,endothelins,nitricoxide(NO), andATP22(Fig.1).
ATP seems tobe the main mediatorin the interaction betweenneuronsandSGC34,35insensoryganglia.P2
recep-torsareexpressedinsensoryneurons(allP2X,exceptP2X7R
andP2YR 1, 2,4, 6),Schwann cells, andSGC(for review see).36Throughcalciumimaging,thepresenceoffunctional
P2YreceptorswasdetectedinGSCofintactTGofrodents.37
Thisexpression wasconfirmedinstudieswithculturedTG cells, and the receptors were classified asP2Y 1, 2, 4 6, 12 and 13 subtypes.18,19 In DSG, only the level of mRNA
wasassessed.38 Among theionotropic receptors,P2X 4 and
P2X739,40 subtypesare found in the SGC,but recent
stud-ieshavereportedthepossibilitythattheyalsoexpressP2X2
andP2X5receptors.41ThedifferentialexpressionofP2X7and
P2X3receptorsinSGCandneurons,respectively,provideda
waytodistinguishtheactionsofATPinneuronsandSGC.39,40
ItwasfoundthatblockingtheactivationoftheP2X7
recep-torwithanantagonistorreducingitsexpressionusingRNA interference(RNAi)innormalrodents,thereisanincreased neuronalexpressionofP2X3,suggestingthatthetonic
acti-vationoftheSGCP2X7receptorsexertsaninhibitingcontrol
overtheP2X3.Ontheotherhand,theATPreleasedby
neu-ronscan activatetheP2X7 receptoroftheSGCleadingto
the release of cytokines, namely TNF-␣, which enhances theneuronP2X3 receptormediatedresponse.Thus,itwas
concludedthattheP2X7exertsinfluence,eitherexcitatory
or inhibitory, onthe sensoryneuronsoma.20,40 It wasalso
shownthatthevesicularreleaseofATPbythecellbodyof neuronsintheDSGactsontheP2X7receptorbyincreasing the intracellular concentration of Ca2+ in the surrounding
CGRP1 NK1
P2
GC cGMP
IL-1β
TNFα
IL-1RI
Depolarization
Neuron Activation
TNF-αRI No
ATP Substance
CGRPP
Satellite glial cells
↑
Figure 1 Intraganglionar communication. After peripheral nerveinjuryasomaticneurotransmitterreleaseoccurs,suchas thecalcitoningene-relatedpeptide(CGRP),substanceP, adeno-sinetriphosphate(ATP),andnitricoxide(NO)inperineuronal environment.Thesemediatorsactivatesatelliteglialcellsvia theirreceptorslocatedonthesurfacemembraneofcells.This activationleadstoreleaseofcytokinessuchastumornecrosis factoralpha(TNF␣)andinterleukin1beta(IL-1),whichinturn caninfluenceneuronalexcitability throughspecificreceptors (TNF␣-RIandIL-1RI).AdaptedwithpermissionRef.22.
SGC.ThisfindingisrelevantbecausethewavesofCa2+ are
usedasa mechanismof informationtransmissionnetwork among astrocytes, mediated by ATP and gap junctions.42
Thus,giventhatSGCexpressP2receptorsandareconnected bygapjunctions,itwaspossibletoconcludethatthesecells, similartoastrocytes,wouldalsohavetheabilitytosustain calciumwaves.Inthissense,astudywasconductedin pri-maryculturesofTGanditwasfoundthattheelectricalor mechanical stimulationof asingle cellbody promoted an increaseinintracellularcalciuminneuronsandsurrounding SGCbyapropagationsimilartotheCa2+ waves.This
prop-agationwasessentiallymediatedbyP2receptorsand,toa lesserextent,bythegapjunctions,whichdemonstratedthe existence ofa bidirectionalcommunication between neu-ronsandSGCperformedbyCa2+.35
As forsubstance P,the findingofitsincreasedsomatic release after orofacial inflammation was the first indica-tionthatthissubstanceplaysanimportantroleinparacrine signalingestablishedingangliaafterinflammation.31,43This
non-nociceptive surroundingneurons.44,45 Moreover,it has
beensuggestedthatthisneuropeptidecanactivatetheSGC throughNK1 receptors,sothatSGCrespondwiththe syn-thesis and release of IL1-.22 The NK1 expression in SGC
wasnotdirectlymeasured,buttheNK1expressionwithhigh affinityfor substance Phasbeen shownin astrocytesand microglia.46
ThefindingthatintraganglionarreleaseofCGRP occurs afteractivationoftrigeminalafferentneurons,alongwith theCGRP1receptorexpressioninneuronsandDSG,21made
thisneuropeptideacandidateformediatingtheneuron---SGC interaction.This hypothesiswasreinforcedbyTGstudies, whichhaveshownthatCGRPcanactinanautocrinemanner, stimulating the promoter activity of CGRP and increasing mRNAlevels.47 Furthermore,it mayalsohave aparacrine
action on CGS, regulating the release of cytokines and chemokines48 and increasing the expression of inducible
nitric oxide synthase (iNOS), as well as the release of NO.21 Nitric oxide is also a potential messenger between
neurons and SGC,as NO released by neurons after nerve injuryacts onSGCcausing increasedexpression of guany-late cyclase ␣1, which catalyzes the formation of cyclic guanosinemonophosphate.49Recently,theextreme
sensitiv-ityoftheTGSGCtoendothelin-1byET-Breceptoractivation wasalsodemonstrated.50 Becausesensoryneuronsexpress
endothelin-1mRNA, this peptide may also be one of the actorsinthecommunicationbetweenSGCandtheneuron.51
ThefunctionalexpressionofNMDArinSGC,whose activ-itymayalsomodulatetheSGC---neuroninteractioninDSG,26
wasalsoverifiedrecently.Severalfactsindicatethe occur-rence of glutamate release in ganglion, particularly the presenceofglutamatereceptors52 andvesicularglutamate
transportersintheneuronalbody.53Besides thisevidence,
allproteinsnecessary fortheuptake andrecycling of glu-tamate were found in the SGC, including the glutamate aspartatetransporter(GLAST)andglialglutamaterecycling enzymes,suchasGS.6,54Inastudythatsoughttounderstand
the roleof glutamate in trigeminal ganglia, it was found thatblocking thesynthesis ofSGin SGCleadstoreduced activationthresholdformechanicalstimulationoftheface. Supposedly,thiseffectisassociatedwithdecreasedrelease ofglutaminebySGCandhencetoadecreaseinglutamine availableforuptakebytheneurontoproduceglutamate.55
SGC
response
to
nerve
injury
and
effect
on
nociception
Thestudiesevaluatingtheroleofsensorygangliainchronic painwerefocusedonchangesinsensoryneuronsafternerve injury.Changes in intrinsic properties of the pericardium maylead tohyperexcitability, characterized by increased incidence of spontaneous activity and reduced activation thresholdbyperipheralstimuli,whichresultinhyperalgesia and allodynia phenomena seen after injury.56,57 Research
in an animal model of pain, mostlyperformed in rodents andbasedprimarily onperipherallesionsduetoaxotomy, inflammation or constriction, indicates that nerve injury not only induces changes in neurons but also in sensory ganglia SGC. Similar to the CNS glial cells, the SGC are activated underthese conditions. The concept of activa-tionis basedonthenotionthat,undernormalconditions,
glial cells are spectators of the nociceptive process, but afterperipheralinjury,theyreactexhibitingmorphological changesandreleasingglialmediators.4Becauseneuronsare
thelesiontarget,thechangesseeninSGCaresecondaryto neuronalchangesandinvolveactivationofsignaling mech-anismsbetweenneuronsandthesecells.
Theeventthattriggersthesechangesseemtoberelated toincreasedneuronalfiringinducedbynerveinjury.58This
hypothesis is basedon severalassumptions. First, experi-ments with variouspain models and methods of blocking neuronalactivitydemonstratedthattheblockadeof sponta-neousactivitypreventsthedevelopmentofpainassociated with non-physiological behavior. One of the studies that establish this relationship usedtwodifferent painmodels in rodents: one induced by chronic constriction injury of thesciaticnerveandanotherinducedbyaxotomyand liga-tionoftibialandcommonfibularnerves.Theeffectoftwo action potential blockers (tetrodotoxin and bupivacaine) wastestedintheseanimals,administeredalone,innormal andinjuredmice.Behavioraltestsforthermalhyperalgesia and mechanical allodynia in these same animals showed a reduction in the behavioral signs of pain. In the same study,theseblockersapplicationpreventedthespontaneous activityintheinjuredsciaticnerve,assessedby electrophys-iologyaftertheadministrationofblockers,beforeandafter the animalswere subjectedto nerveinjury.59 The second
assumptionisbasedontheobservationthattheactivationof SGCfromDSGaftersciaticnerveinjuryispreventedbythe localnerveconductionblockade.Inthisstudy,performedin amodelofL4spinalnerveaxotomy,withanimplanted infu-sionpumpoftetrodotoxin,therewasamarkedreductionin theSGCactivation,whendetectedbytheassessmentofthe GFAP levels of expression using immunohistochemistry. In thatsamestudy,thisresultwasconfirmedbylocal applica-tionofothersodiumchannelblockers(bupivacaine)inrats withperipheralinjuryinducedbythebindingofthefibular andtibialnerves.60
Despite this evidence, the abnormalspontaneous neu-ronalactivityisonlyonecandidateforatriggeringevent.In fact,thisissueisatopicofrecentresearch,andthereare stillnostudiesclarifyingthemechanismsresponsibleforthe SGCactivation. However,severalstudies have shown that thesecellsundergodeepchangesinresponsetonerveinjury, mainly characterized by an increased expression of GFAP, decreased expression and sensitivity of potassium chan-nels, increased SGCcoupling by gap junctions, increased sensitivity to ATP, alteredexpression of purinergic recep-tors, and release of ATP and cytokines.6,22,54,61 There is
alsoevidencethatthesechangesmaycontributetochronic pain.62,63
Increased
expression
of
GFAP
UndernormalconditionstheSGCexhibitlow,almost unde-tectable,levelsofGFAPbyimmunohistochemistry,bywhich theobservationinseveralstudiesofitsincreasedexpression after injury made it the essential marker in the assess-ment ofSGCactivation.16,60,62,64---68 The significanceof this
via glutamate. Accordingtothis hypothesis, the extracel-lular increase of this neurotransmitter would trigger the GFAP increase required to support the increased expres-sion of GLAST, since this filament is essential to anchor GLAST tothe plasma membrane of astrocytes.69,70 Based
on these findings, it was hypothesized that, as in astro-cytes, the glutamate released in sensory ganglia could trigger the increased expression of GFAP in SGC, but so farthis hasnotbeen proven.54 Even so,theexpressionof
GFAP is considered the best known marker of SGC acti-vation. Its usefulness could be proven in a study that used a model of neuropathic pain induced by ligation of the fifth lumbar spinal nerve, in which the GFAP reac-tivity to assess the SGC activation was analyzed. It was demonstratedthatthisactivationcontributestothe main-tenance of the neuropathic pain symptoms in the early periodofdisease,morepreciselytheonsetofmechanical allodynia.58
Increased
expression
and
sensitivity
of
potassium
channels
SGC are responsible for perineural K+ homeostasis
regu-lated throughthe input channels of rectified K+ currents’
glialcell-specific(Kir4.1)15,16,71 andgap junctions.52,72 The
conventional model of neuronal ionic balance predicts that increased K+ levels correspond to increased
neu-ron excitability, which can lead to changes in sensory perception.73 Thus, maintenance of low extracellular K+
concentrations,mediatedbySGC,maybecrucialtocontrol therestingmembranepotentialandneuronalexcitability.15
To help answer this question, several studies have eval-uated if the SGC response to nerve injury would involve changesinKirchannels.Thus,thedecreasedexpressionof Kir4.1 was observed in SGC of TG, in a model of chronic constrictionoftheinfraorbitalnerve.15,74Also,invitro
elec-trophysiological studiesof DRGfromanimalssubjected to chronic compression of these ganglia, it was found that theSGCininjuredgangliaexhibitedasignificantreduction of currents mediated by Kir.16 Similar results were found
whentheeffectofperipheralinflammation(facialskin)on currents mediated by Kir in rats in vivo was investigated by immunohistochemistry and patch clamp. In this study there wasa significantly smaller increase of the currents mediated by Kir in rats with inflammation, compared to naivemice,followedbyadecreaseintheactivation thresh-oldtomechanicalstimuli,suggestiveofhyperalgesia.75The
aforementioned studies showed that the SGCresponse to different types of injury includes the decreased expres-sionof theKir channels anddecreasedrectifying currents mediated bythem. On the other hand, inthe absenceof injury,decreased expressionofKirchannelshasanimpact onneuronalactivity,asshown bythe specificsilencingof Kir4.1 expression using RNAi. This silencingwas sufficient toproducebehavioralsignsofpaininmice,characterized bythedevelopmentofspontaneous(measuredbyincreased frequencyofclosingtheeyes)andevokedpain(facial allo-dynia), which reinforced the importance of SGC in the clearanceofK+anditsabilitytopromotechangesin
neu-ronalactivity.74
Increased
coupling
between
SGC
via
gap
junctions
ThebridgingconnectionbetweenSGCdistinctunitsandthe increasednumberofgapjunctionsbetweenthemwerethe first evidence that SGC change in response to peripheral nerveinjury.12,13Subsequently,studiesofelectrophysiology
anddye injectionconfirmed this increased couplingafter anerveinjury.15,76Indeed,theincreaseddensity(number)
ofgapjunctionsandcouplingbetweensensorygangliaSGC afternerveinjuryareaconsistentfindinginseveralstudies of pain.In SGC of DSG,this change was found in several models of pain, from colon63,76 and thigh71 inflammation
tosciaticnerveneuritis72 and DSGchronic compression.17
TG studies also showed increased coupling between SGC inmodels of orofacial pain,particularly after infraorbital nerveaxotomy15orchronicconstriction.16Inordertoassess
the significance of this increase in the coupling between theSGC,found in many modelsofchronic pain, apotent gapjunctionblocker(carbenoxolone)wasused,which sup-pressedtheincreasedcouplingbetweentheSGCcausedby an inflammation previously induced by injection of com-plete Freund’s adjuvant in the thigh, with an increase in the threshold of activation stimuli.71 Similar analgesic
effectswere seen withother gap junctionblockers, such asmeclofenamic and palmitoleic acids, which favors the hypothesisthatgapjunctionsbetweenSGCplayan impor-tantroleinneuronalexcitability.63
The identification of a constituent molecule of gap junctions, particularly connexin43 (Cx43), allowed a dif-ferent experimental approach to study the role of these junctionsinSGC.Thus,itwasfoundthatafterthe infraor-bitalnerveinjury,anincreasedexpressionofconnexinalso occurs.16,55,62UsingRNAiforCx43tochangethegapjunction
properties,it wasobservedthata disturbanceinthis pro-teinexpressionisenoughtocausechangesintheactivation thresholdofafferentneurons.16Furthermore,theinhibition
ofCx43expressioninTGofmicewithorofacialneuropathic paininducedbychronicconstrictionoftheinfraorbitalnerve wasaccompaniedby adecreasedbehaviorof spontaneous andevokedpain.62 Ontheotherhand,whentheinhibition
was carried out in the trigeminal ganglia of naive mice, a nociceptiveresponse identical tothat seen after nerve damagehasoccurred.16,62 Thesestudies suggest that
inhi-bitionofCx43canhavepro-nociceptiveorantinociceptive effectin normal animals after nerve injury.In trigeminal ganglia,some studies have found increased expression of other types of connexins, namely Cx36 and Cx40, after temporomandibularjointinflammation,suggestingthatthe typeofconnexinwhoseexpressionincreasesdependsonthe painmodelinquestion.73 Regardlessoftheapproach,
evi-denceindicatesthatthecouplingbetweentheSGC,which involvesgap junctions and consequently Cx43,appears to beassociated withtheneuropathic paindevelopmentand maintenance.Theunderlyingmechanismsarestillunknown, butseveralhypotheseshavebeenidentified,namelytherole ofthiscouplinginmaintainingtheelectrochemicalgradient andbufferingof K+ toallowthe rapidredistribution ofK+
afternerveinjury.15Otherhypothesessuggestthatthis
allogeneicsubstances(e.g.,ATP,Ca2+)fromthesiteofinjury
toadjacentareas,leadingtoanamplificationoftheprimary injury.63Finally,otherssuggestthatitwillhaveanactionin
therecyclingofglutamate.62
Increased
sensitivity
to
ATP
and
altered
expression
of
purinergic
receptors
Several studies reported the plasticity of P2 receptors on SGC in response to nerve damage or inflammation, which results in an increased sensitivity to ATP and alteredexpressionofthesereceptors.18,41,74Withtheuseof
microfluorometrytodeterminethecytosolicconcentration ofCa2+,therewasanincreasedsensitivitytoATPincultured
TGof SGCofmice withinduced facialskin inflammation. SimilarresultswerefoundinintactTGanalysisinvitro,from ratssubjectedtoaxotomyoftheinfraorbitalnerve,inwhich a100-foldincreaseinsensitivityofSGCtoATPwasrecorded. Moreover,theoccurrenceofareversalinpurinergic recep-torsubtypes in cells of TG culture can be observed with theuseofpharmacologicaltools. Infact,innormalmice, theresponsetoATPwasmediatedbyP2Yreceptors,while inmicewithinflammation,itwaspredominantlymediated byP2X.41Recently,apreliminarymodeloftheSGCrolein
chronicpainhasbeenproposedinanattempttoexplainhow theincreasedgapjunctionsandhighsensitivitytoATPcan leadtoabnormalneuronalactivitybothininjuredand non-injuredneurons.Thismodelisbasedontheknowledgethat whenaperipheralnerveinjuryincreasestheexcitabilityof sensoryneurons,italsoincreasestheexcitatorysignalsfrom injuredneurons tothe SGCthat surroundthem, andthat thesecells,communicatingwithSGCofadjacentunits,will influenceitsneuron.61 Italsopredictsthat inresponse to
peripheralinjury asomaticATPreleaseoccurs,whichwill activatetheP2receptorsonthesurroundingSGCand neu-ronitself.Inturn,thisactivationshouldleadtoanincrease inintracellularCa2+inbothcelltypesandconsequentlyATP
releasefrombothneuronsandSGC(whoselevelof sensitiv-itytoCa2+ increasesafterinjury).ThisATPincrease,along
withtheincreasednumbersofgapjunctions betweenSGC ofneighboring perineuralsheaths,willallow the propaga-tionofCa2+ wavestotheseSGCandneighboring neurons,
influencingtheexcitabilityofneuronsnotdirectlyaffected bythelesion(Figure2).Thisintraganglionarcommunication modelcouldbeoneexplanationofhowaperipheralinjury canaffectalargenumberofsensoryneurons,contributing tosignalpropagationandchronicpain.35,41
Production
of
cytokines
SGChavesimilarcharacteristicstocellsoftheimmune sys-tem, beingactivated by monocyte chemotacticprotein-1 (MCP-1)throughthereceiver,andproducingcytokinessuch asTNF-␣,23,67IL-177andIL6.78TheSGCabilitytosynthesize
TNF-␣inresponsetoaperipheralinjurywasshowninone adaptedmodeloflumbarspinefacetjointinjuryinmice67
andin three painmodels in sciatic nerve(unilateral par-tialconnection,spinalnerveconnection,andtransaction). In these models, immunocytochemistry showed increased expressionofTNF-␣andTNF-␣-1receptoronneuronsand itsSGC.23 Moreover,itwasfoundthattheTNF-␣ activates
ATP SGC Peripheral
injury
Neuron 1
Gap junction
SGC
ATP
Neuron 2
↑[Ca2+]
↑[Ca2+]
ATP
Figure2 Modelofinteractionbetweenneuronsviasatellite glialcells.Peripheralnerveinjuryleadstoreleaseofsomatic ATP thatacts viapurinergic receptors onsatellite glial cells (SGC),leadingtoasignificantincreaseinintracellularcalcium concentration([Ca2+])inthesecells.Thecommunicationwith
satellite glial cells ofneighboring perineural sheathand the wavepropagationofCa2+tothesecellsoccursthroughgap
junc-tions,whichleadstothereleaseofATPbytheseneighboring satelliteglialcells.ThisATPbindstotheneuronalpurinergic receptors,influencingtheexcitabilityoftheneuron thatwas notdirectlyaffectedbytheinjury.
theSGCcausingincreasedphosphorylationofproteinkinase regulatedbyextracellularsignals(ERK79).Interestingly,the
long-termincreasedactivationofthisproteininSGCafter nerveinjuryhasbeenassociatedwithchronicpain.80
AnothercytokineproducedbyactivatedSGCisIL-1.The roleofinterleukininthemechanismunderlyingthe devel-opmentof hyperalgesiaandallodyniafollowingperipheral inflammation has been extensively investigated in animal models of cutaneous inflammation. For example, it was shownthatSGCrespondtothisinflammationwithincreased production of IL-1 and that an increased expression of IL-1RI inneurons occurssimultaneously.It wasalsoshown thattheapplicationofIL-1inneuronsproducesagreater increase of the action potential firing rate on inflamed mice compared to normal mice. Thus, it was postulated that SGCcan modulatethe excitability of TGnociceptive neurons via IL-1, inducingmembrane depolarization and increased expression of IL-1RI in neuronal body.77 This
orofacial inflammatory pain in vitro, it was found that IL-1- suppresses the K+ currents of voltage-dependent
channels in small diameter neurons (i.e., mostly A␦ and C nociceptive neurons). These data suggest that IL-1-
released by SGCactivated afterinflammation potentiates theexcitabilityofnociceptiveneuronsbysuppressionofK+
currents.82Gatheredevidenceallowedtheconstructionofa
mechanismunderlyingtheinflammatoryhyperalgesiawhich predictsthat,ininflammatoryconditions,theactivationof SGCcanincreasetheexcitabilityofA␦nociceptiveneurons via IL-1. According to this hypothesis, the substance P releasedfromthecellbodyofprimaryafferentnociceptive neurons, activated by peripheral injury/inflammation,31
willactonNK1receptorsandwillsomehowpotentiatethe synthesisand/orreleaseofIL-1bySGC.Thiscytokinewill, inturn,suppresstheneuronvoltage-dependentK+channels,
thus contributing to the central sensitization responsible forthehyperalgesiaandallodyniaafterinflammation.22
Finally, IL-6 also appears to be involved in the SGC responsetoneuroinflammation.Infact,abilateralincreased expressionofIL-6wasobservedinSGCofDSGaswellasinits receptorintheipsilateralganglion,aftersciaticnerveinjury by chronic constriction.78 In short,cytokines areinvolved
intheinteractionbetweenaneuronandSGC,andthereis growingevidenceof apossibleroleforcytokines originat-inginsensorygangliaintheinductionandmaintenanceof neuropathicpain.83
Final
considerations
The progress in understanding the SGC biology, and the recognitionof itsinteractionwithsensoryneurons,called the attention of the scientific community to the role of these cells in the nociceptive process. The perineu-ronal environment monitoring exerted by SGC as well as the cell communication occurring in sensory ganglia (neuron---neuron, neuron---SGC, SGC---SGC) can affect neu-ronalexcitability.22,35,61Infact,thechangesresultingfrom
the activation of SGC, which have been observed in dif-ferent pain models, enabled the observation that these cellsmaymodulatechronicpain.16,22,54,63,71Therefore,
con-trarytowhatwasinitiallypostulated,sensoryganglionmay becomethefirstlevelofthepathophysiologicalchangesof modulationofafferentsignaling,asitallowstheinteraction betweendifferenttypesofinformationandseemstobea triggerofthecentralsensitizationmechanismofthespinal corddorsalhornneurons.22Thus,knowledgeabouttheSGC
anditsmechanismsof interactionwiththe neuronalbody assumesagrowingimportanceinthesearchfornewtargets forchronicpaintreatment.
Conflicts
of
interest
Theauthorsdeclarenoconflictsofinterest.
Acknowledgements
The authors acknowledge the help given by Prof. Daniel Humberto Pozza, Department of Experimental Biology,
Faculdade deMedicine doPorto, in reviewing thetext to PortugueseofBrazil.
References
1.MerskeyH,LindblomU,MumfordJM,NathanPW,Sunderland S.PartIIIPainterms:acurrentlistwithdefinitionsandnotes onusage.In: MerskeyH,Bogduk N,editors.Classificationof chronicpain.2nded.IASP;1994.
2.McMahonSB,KoltzenburgM,editors.WallandMelzack’s--- text-bookofpain.5thed.Philadelphia:Elsevier;2006.
3.ScholzJ,WoolfCJ.Canweconquerpain?NatNeurosci.2002;5 Suppl.:1062---7.
4.McMahonSB,MalcangioM.Currentchallengesinglia-pain biol-ogy.Neuron.2009;64:46---54.
5.HananiM.Satelliteglialcells:morethanjust‘ringsaroundthe neuron’.NeuronGliaBiol.2010;6:1---2.
6.HananiM.Satelliteglialcellsinsensoryganglia:fromformto function.BrainResBrainResRev.2005;48:457---76.
7.JessenKR,MirskyR.Theoriginanddevelopmentofglialcells inperipheralnerves.NatRevNeurosci.2005;6:671---82. 8.PanneseE.Thesatellitecellsofthesensoryganglia.AdvAnat
EmbryolCellBiol.1981;65:1---111.
9.BungeMB,BungeRP,PetersonER,etal.Alightandelectron microscopestudyoflong-termorganizedculturesofratdorsal rootganglia.JCellBiol.1967;32:439---66.
10.PanneseE.Thestructureoftheperineuronialsheathof satel-lite glial cells (SGCs) in sensory ganglia. Neuron Glia Biol. 2010;6:3---10.
11.Pannese E,LeddaM, ArcidiaconoG, etal. Clustersofnerve cellbodiesenclosedwithinacommonconnectivetissue enve-lopeinthespinalgangliaofthelizardandrat.CellTissueRes. 1991;264:209---14.
12.Hanani M, Huang TY, Cherkas PS, et al. Glial cell plasticity in sensory ganglia induced by nerve damage. Neuroscience. 2002;114:279---83.
13.Pannese E,Ledda M, Cherkas PS, et al. Satellite cell reac-tionsto axoninjury ofsensoryganglionneurons:increasein emumberofgapjunctionsandformationofbridgesconnecting previouslyseparateperineuronialsheaths.AnatEmbryol(Berl). 2003;206:337---47.
14.PanneseE,ProcacciP,LeddaM,etal.Age-relatedreductionof thesatellitecellsheatharoundspinalganglionneuronsinthe rabbit.JNeurocytol.1996;25:137---46.
15.CherkasPS,HuangTY,PannickeT,etal.Theeffectsofaxotomy onneuronsandsatelliteglialcellsinmousetrigeminalganglion. Pain.2004;110:290---8.
16.VitJP,JasminL,BhargavaA,etal.Satelliteglialcellsinthe trigeminalganglionasadeterminantoforofacialneuropathic pain.NeuronGliaBiol.2006;2:247---57.
17.Zhang H, Mei X, Zhang P, et al. Altered functional proper-tiesofsatelliteglialcellsincompressedspinalganglia.Glia. 2009;57:1588---99.
18.CerutiS,FumagalliM,VillaG,etal.Purinoceptor-mediated cal-ciumsignalinginprimaryneuron-gliatrigeminalcultures.Cell Calcium.2008;43:576---90.
19.VillaG,FumagalliM,VerderioC,etal.Expressionand contribu-tionofsatelliteglialcellspurinoceptorstopaintransmission in sensory ganglia: an update. Neuron Glia Biol. 2010;6: 31---42.
20.ZhangX,ChenY,WangC,etal.NeuronialsomaticATPrelease triggersneuron-satelliteglialcellcommunicationindorsalroot ganglia.ProcNatlAcadSciUSA.2007;104:9864---9.
22.TakedaM,TakahashiM,MatsumotoS.Contributionofthe acti-vationofsatellitegliainsensorygangliatopathologicalpain. NeurosciBiobehavRev.2009;33:784---92.
23.DubovyP,JancalekR,KlusakovaI,etal.Intra-and extraneuro-nialchangesofimmunofluorescencestainingforTNF-alphaand TNFR1inthedorsalrootgangliaofratperipheralneuropathic painmodels.CellMolNeurobiol.2006;26:1205---17.
24.LiM,ShiJ,TangJR,etal.EffectsofcompleteFreund’sadjuvant onimmunohistochemicaldistributionofIL-1betaandIL-1RIin neuronsandgliacellsofdorsalrootganglion.ActaPharmacol Sin.2005;26:192---8.
25.PomonisJD,RogersSD,PetersCM,etal.Expressionand localiza-tionofendothelinreceptors:implicationsfortheinvolvement of peripheral glia in nociception. J Neurosci. 2001;21: 999---1006.
26.CastilloC,NorciniM,MartinHernandezLA,etal.Satelliteglia cellsindorsalrootgangliaexpressfunctionalNMDAreceptors. Neuroscience.2013;240C:135---46.
27.ShinderV,DevorM.Structuralbasisofneuron-to-neuron cross-excitationindorsalrootganglia.JNeurocytol.1994;23:515---31. 28.AmirR,DevorM.Chemicallymediatedcross-excitationinrat
dorsalrootganglia.JNeurosci.1996;16:4733---41.
29.Amir R, DevorM.Functional cross-excitation between affer-ent A- and C-neurons in dorsal root ganglia. Neuroscience. 2000;95:189---95.
30.HuangLY,NeherE.Ca(2+)-dependentexocytosisinthesomata ofdorsalrootganglionneurons.Neuron.1996;17:135---45. 31.MatsukaY,NeubertJK,MaidmentNT,etal.Concurrentrelease
ofATPandsubstancePwithinguineapigtrigeminalgangliain vivo.BrainRes.2001;915:248---55.
32.Hayasaki H, Sohma Y, Kanbara K, et al. A local GABAergic system within rat trigeminal ganglioncells. Eur J Neurosci. 2006;23:745---57.
33.McCarthyPW,LawsonSN.Differingactionpotentialshapesin ratdorsalrootganglionneuronesrelatedtotheirsubstanceP andcalcitoningene-relatedpeptideimmunoreactivity.JComp Neurol.1997;388:541---9.
34.GuY,ChenY,ZhangX,etal.Neuronialsoma-satelliteglialcell interactionsinsensorygangliaandtheparticipationof puriner-gicreceptors.NeuronGliaBiol.2010;6:53---62.
35.Suadicani SO, Cherkas PS,Zuckerman J, et al. Bidirectional calcium signaling between satellite glial cells and neurons in cultured mouse trigeminal ganglia. Neuron Glia Biol. 2010;6:43---51.
36.BurnstockG.Physiologyandpathophysiologyofpurinergic neu-rotransmission.PhysiolRev.2007;87:659---797.
37.WeickM,CherkasPS,Hartig W,et al.P2 receptorsin satel-lite glial cells in trigeminal ganglia of mice. Neuroscience. 2003;120:969---77.
38.KobayashiK,FukuokaT,YamanakaH,etal.Neuronsandglial cellsdifferentiallyexpressP2YreceptormRNAsintheratdorsal rootganglionandspinalcord.JCompNeurol.2006;498:443---54. 39.Kobayashi K, Fukuoka T, Yamanaka H, et al. Differential expressionpatternsofmRNAsforP2Xreceptorsubunitsin neu-rochemicallycharacterizeddorsalrootganglionneuronsinthe rat.JCompNeurol.2005;481:377---90.
40.ChenY,ZhangX,WangC,etal.ActivationofP2X7receptors inglialsatellitecellsreducespainthroughdownregulationof P2X3receptorsinnociceptiveneurons.ProcNatlAcadSciUS A.2008;105:16773---8.
41.Kushnir R, Cherkas PS, Hanani M. Peripheral inflammation upregulatesP2Xreceptorexpressioninsatelliteglialcellsof mouse trigeminal ganglia: a calcium imaging study. Neuro-pharmacology.2011;61:739---46.
42.ScemesE,GiaumeC.Astrocytecalciumwaves:whattheyare andwhattheydo.Glia.2006;54:716---25.
43.Neubert JK, Maidment NT, Matsuka Y, et al. Inflammation-induced changes in primary afferent-evoked release of
substance P within trigeminal ganglia in vivo. Brain Res. 2000;871:181---91.
44.TakedaM,TanimotoT,IkedaM,etal.Temporomandibularjoint inflammation potentiates the excitability of trigeminal root ganglionneuronsinnervating thefacialskininrats.J Neuro-physiol.2005;93:2723---38.
45.TakedaM,TanimotoT,NasuM,etal.ActivationofNK1 recep-tor of trigeminal root ganglion via substance P paracrine mechanismcontributestothemechanicalallodyniainthe tem-poromandibular joint inflammation in rats. Pain. 2005;116: 375---85.
46.MarriottI.The roleoftachykinins incentralnervous system inflammatoryresponses.FrontBiosci.2004;9:2153---65. 47.ZhangZ, Winborn CS, Marquezde Prado B, etal.
Sensitiza-tionofcalcitoningene-relatedpeptidereceptorsbyreceptor activity-modifyingprotein-1inthetrigeminalganglion.J Neu-rosci.2007;27:2693---703.
48.ThalakotiS,PatilVV,DamodaramS,etal.Neuron-gliasignaling in trigeminal ganglion: implications for migraine pathology. Headache.2007;47:1008---23,discussion1024---5.
49.ThippeswamyT,MorrisR.Therolesofnitricoxideindorsalroot ganglionneurons.AnnNYAcadSci.2002;962:103---10. 50.Feldman-GoriachnikR,HananiM.Functionalstudyof
endothe-lin B receptors in satellite glial cells in trigeminal ganglia. Neuroreport.2011;22:465---9.
51.Giaid A, Gibson SJ, Ibrahim BN, et al. Endothelin 1, an endothelium-derivedpeptide,is expressedinneurons ofthe humanspinalcordanddorsalrootganglia.ProcNatlAcadSciU SA.1989;86:7634---8.
52.WillcocksonH,ValtschanoffJ.AMPAandNMDAglutamate recep-torsarefoundinbothpeptidergicandnon-peptidergicprimary afferentneuronsintherat.CellTissueRes.2008;334:17---23. 53.BrumovskyP,WatanabeM,HokfeltT.Expressionofthe
vesicu-larglutamatetransporters-1and-2inadultmousedorsalroot gangliaand spinalcordand theirregulationbynerveinjury. Neuroscience.2007;147:469---90.
54.OharaPT,VitJP,BhargavaA,etal.Gliopathicpain:when satel-liteglialcellsgobad.Neuroscientist.2009;15:450---63. 55.JasminL, Vit JP, Bhargava A, et al. Can satellite glialcells
be therapeutic targets for pain control? Neuron Glia Biol. 2010;6:63---71.
56.AmirR,MichaelisM,DevorM.Membranepotentialoscillations indorsalrootganglionneurons:roleinnormalelectrogenesis andneuropathicpain.JNeurosci.1999;19:8589---96.
57.Zimmermann M. Pathobiology of neuropathic pain. Eur J Pharmacol.2001;429:23---37.
58.LiuFY,SunYN,WangFT,etal.Activationofsatelliteglialcells inlumbardorsalrootgangliacontributestoneuropathicpain afterspinalnerveligation.BrainRes.2012;1427:65---77. 59.XieW,StrongJA,MeijJT,etal.Neuropathicpain:early
spon-taneousafferentactivityisthetrigger.Pain.2005;116:243---56. 60.XieW,StrongJA,ZhangJM.Earlyblockadeofinjuredprimary sensoryafferentsreducesglialcellactivationintworat neuro-pathicpainmodels.Neuroscience.2009;160:847---57.
61.HananiM. Intercellularcommunication insensory gangliaby purinergicreceptorsandgapjunctions:implicationsforchronic pain.BrainRes.2012;1487:183---91.
62.OharaPT,VitJP,BhargavaA,etal.Evidenceforaroleof con-nexin43intrigeminalpainusing RNAinterferenceinvivo.J Neurophysiol.2008;100:3064---73.
63.HuangTY,BelzerV,HananiM.Gapjunctionsindorsalroot gan-glia:possiblecontributiontovisceralpain.EurJPain.2010;14, 49.e1---11.
64.StephensonJL,ByersMR.GFAPimmunoreactivityintrigeminal ganglionsatellitecellsaftertoothinjury inrats.ExpNeurol. 1995;131:11---22.
after inferior alveolar nerve crush in the adult rat. Pain. 1997;73:141---9.
66.OhtoriS,TakahashiK,MoriyaH,etal.TNF-alphaandTNF-alpha receptortype1upregulationingliaandneuronsafterperipheral nerveinjury:studiesinmurineDRGandspinalcord.Spine(Phila Pa1976).2004;29:1082---8.
67.MiyagiM,OhtoriS,IshikawaT,etal.Up-regulationofTNFalpha inDRGsatellitecellsfollowinglumbarfacetjointinjuryinrats. EurSpineJ.2006;15:953---8.
68.SiemionowK, KlimczakA, Brzezicki G, et al.The effects of inflammation on glial fibrillary acidic protein expression in satellitecellsofthedorsalrootganglion.Spine(PhilaPa1976). 2009;34:1631---7.
69.SullivanSM,LeeA,BjorkmanST,etal.Cytoskeletalanchoringof GLASTdeterminessusceptibilitytobraindamage:anidentified roleforGFAP.JBiolChem.2007;282:29414---23.
70.RomaoLF,Sousa VdeO,NetoVM, etal. Glutamateactivates GFAPgene promoter from cultured astrocytesthrough TGF-beta1pathways.JNeurochem.2008;106:746---56.
71.DublinP,HananiM.Satelliteglialcellsinsensoryganglia:their possiblecontributiontoinflammatorypain.BrainBehavImmun. 2007;21:592---8.
72.LeddaM,BlumE,DePaloS,etal.Augmentationingap junction-mediatedcellcouplingindorsalrootgangliafollowingsciatic nerveneuritisinthemouse.Neuroscience.2009;164:1538---45. 73.Garrett FG, Durham PL. Differential expression of connex-insintrigeminal ganglionneurons and satelliteglial cells in responsetochronicoracutejointinflammation.NeuronGlia Biol.2008;4:295---306.
74.ChessellIP,HatcherJP,BountraC,etal.DisruptionoftheP2X7 purinoceptorgeneabolisheschronicinflammatoryand neuro-pathicpain.Pain.2005;114:386---96.
75.LaMotte RH,MaC.Hyperexcitable neuronsand altered non-neuronialcellsinthecompressedspinalganglion.ShengLiXue Bao.2008;60:597---602.
76.HuangTY,CherkasPS,RosenthalDW,etal.Dyecouplingamong satelliteglialcellsinmammaliandorsalrootganglia.BrainRes. 2005;1036:42---9.
77.Takeda M, Tanimoto T, Kadoi J, et al. Enhanced excitabil-ity of nociceptive trigeminal ganglion neurons by satel-lite glial cytokine following peripheral inflammation. Pain. 2007;129:155---66.
78.DubovyP,KlusakovaI,SvizenskaI,et al.Satellite glialcells expressIL-6andcorrespondingsignal-transducingreceptorsin thedorsalrootgangliaofratneuropathicpainmodel.Neuron GliaBiol.2010;6:73---83.
79.Takahashi N, Kikuchi S, Shubayev VI, et al. TNF-alpha and phosphorylation ofERK inDRG andspinal cord:insights into mechanismsofsciatica.Spine(PhilaPa1976).2006;31:523---9. 80.Doya H, Ohtori S, Takahashi K, et al. Extracellular
signal-regulatedkinasemitogen-activatedproteinkinaseactivationin thedorsalrootganglion(DRG)andspinalcordafterDRGinjury inrats.Spine(PhilaPa1976).2005;30:2252---6.
81.Takeda M, Takahashi M, Matsumoto S. Contribution of acti-vated interleukin receptors in trigeminal ganglion neurons to hyperalgesiaviasatellite glialinterleukin-1betaparacrine mechanism.BrainBehavImmun.2008;22:1016---23.
82.Takeda M, Kitagawa J, Takahashi M, et al. Activation of interleukin-1betareceptorsuppressesthevoltage-gated potas-siumcurrentsinthesmall-diametertrigeminalganglionneurons followingperipheralinflammation.Pain.2008;139:594---602. 83.White FA, Jung H, Miller RJ. Chemokines and the